InteractiveFly: GeneBrief
dumpy: Biological Overview | References
| Gene name - dumpy
Synonyms - Cytological map position - 24F4-25A1 Function - extracellular matrix component Keywords - Dpy and Piopio establishing an external scaffold that helps to shape the corneal lens - Dusky-like controls the arrangement of Dumpy and Piopio - Loss of either dusky-like or dumpy delays chitin accumulation and disrupts the outer surface of the corneal lens - artificially inducing apical constriction by activating myosin contraction is sufficient to similarly alter chitin deposition and corneal lens morphology - Dumpy guides the position and direction of buckling-induced folds in wing morphogenesis - Piopio interacts and cooperates with Dumpy at tracheal cells - To resist shear stresses which arise during tube expansion, Piopio undergoes ectodomain shedding by the Matriptase homolog Notopleural, which releases Piopio-Dumpy-mediated linkages between membranes and extracellular matrix |
Symbol - dpy
FlyBase ID: FBgn0053196 Genetic map position - chr2L:4,477,462-4,591,963 NCBI classification - EGF_CA: Calcium-binding EGF-like domain and other motifs Cellular location - a membrane-anchored extracellular protein |
The Drosophila corneal lens is entirely composed of chitin and other apical extracellular matrix components, and it is not known how it acquires the biconvex shape that enables it to focus light onto the retina. This study shows that the zona pellucida domain-containing protein Dusky-like is essential for normal corneal lens morphogenesis. Dusky-like transiently localizes to the expanded apical surfaces of the corneal lens-secreting cells and prevents them from undergoing apical constriction and apicobasal contraction. Dusky-like also controls the arrangement of two other zona pellucida domain proteins, Dumpy and Piopio, external to the developing corneal lens. Loss of either dusky-like or dumpy delays chitin accumulation and disrupts the outer surface of the corneal lens. Artificially inducing apical constriction by activating myosin contraction is sufficient to similarly alter chitin deposition and corneal lens morphology. These results demonstrate the importance of cell shape in controlling the morphogenesis of overlying apical extracellular matrix structures such as the corneal lens (Ghosh, 2024).
Forming the precisely curved architecture of the Drosophila corneal lens from apical ECM (aECM) requires the ZP-domain protein Dyl. Dyl acts transiently at a critical point in development to maintain the apical expansion of corneal lens-secreting cells and to assemble a scaffold containing ZP-domain proteins that acts as a convex outer boundary within which chitin and other corneal lens components are retained. These results add to a growing body of evidence that the shapes of rigid structures composed of aECM rely on specific sites of attachment to the underlying cells that are mediated by ZP-domain proteins (Ghosh, 2024).
The data show that the marked apical expansion of primary pigment cells in the pupal retina is essential to build a foundation for corneal lens assembly. Reversing this expansion either by removing Dyl or by activating myosin contraction results in severe corneal lens defects. The apicobasal contraction that occurs in dyl mutant or MlckCT-expressing ommatidia places the apical surfaces of cone and primary pigment cells in a more basal position than their wild-type neighbors. If secondary and tertiary pigment cells retain their attachments to the aECM, the ommatidial surface could take on a concave shape, potentially explaining the deeper inner curvature of the overlying corneal lenses. The smaller surface area of the central cells could also directly alter the shape of the corneal lens if this structure is assembled one layer at a time, like the tectorial membrane. Such a pattern of assembly would be consistent with labeling of the outer surface of the early corneal lens and the inner surface of the adult corneal lens by the chitin-binding probe, which may preferentially bind to newly deposited chitin. Constriction of the apical cell surface has been shown to produce ridges of aECM in the C. elegans cuticle and the Drosophila trachea; the results suggest that apical expansion can also drive the morphogenesis of specific aECM structures. However, not all aECM remodeling requires cell shape changes; for example, formation of sex-specific pores that accommodate chemosensory cilia in the C. elegans cuticle appears to depend on gene expression in the underlying glia rather than on the shape of these cells (Ghosh, 2024).
The notable apical constriction of cone and primary pigment cells that was observed in dyl mutants implies that Dyl is required to maintain their expanded shape. Dyl is located on the apical surfaces of these cells, and it and other ZP-domain proteins have been shown to attach the plasma membrane to the aECM. These attachments can alter cell shape; for instance, apical expansion of cells in the pupal wing is dependent on the ZP-domain proteins Miniature and Dusky; NOAH-1 and NOAH-2 are required for the concerted cell shape changes that drive elongation of the C. elegans embryo , and Hensin is needed to convert flat β- to columnar α-intercalated cells in the kidney. The NOAH-1, NOAH-2, and FBN-1 ZP-domain proteins in the embryonic sheath of C. elegans resist deformation of the epidermis by mechanical forces. It is suggested that attachment to the aECM by Dyl enables cone and primary pigment cells to resist mechanical tension exerted by the actomyosin cytoskeleton. Active myosin and βH-spectrin accumulate in dyl mutant cells, and reducing the level of these components partially rescues apical constriction. However, these proteins appear disorganized and do not form foci like those seen in the wild-type cells. Similar foci are associated with changes in apical cell area driven by pulsatile constriction and ratcheting, but continuous constriction can occur with a diffuse apical actomyosin distribution, for instance, in boundary larval epithelial cells. The loss of apical anchorage may cause dyl mutant cells to constrict in a continuous, passive manner that does not involve controlled ratcheting (Ghosh, 2024).
Although electron micrographs suggest that Dyl establishes a physical link between the apical plasma membrane and the aECM, this role at contact sites is surprising because it is likely that the extracellular domain of Dyl is proteolytically cleaved from its transmembrane domain. Overexpressed Dyl appears to diffuse away from the cells that express it (Adler, 2013; Nagaraj, 2012), and Dyl contains a putative furin cleavage site that has been used to enable secretion of a reporter protein. Nevertheless, loss of dyl has cell-autonomous effects on apical constriction and chitin accumulation in the retina, indicating that the protein does not freely diffuse in this tissue and may associate with other transmembrane proteins and/or with immediately overlying aECM. Alternative explanations for Dyl function are also possible. Rather than directly attaching the membrane to the matrix, Dyl might affect the properties of the developing aECM, for instance, by endowing it with sufficient tensile strength to counteract cytoskeletal forces that would otherwise drive apical constriction of the cone and pigment cells (Ghosh, 2024).
The data also reveal a role for other ZP-domain proteins, including Dpy and Pio, in establishing an external scaffold that helps to shape the corneal lens. Although dpy mutant ommatidia show little constriction, indicating that dpy acts downstream of or in parallel to the cell shape changes, they fail to accumulate chitin at the normal time and produce deformed corneal lenses. As chitin is not found trapped inside dyl or dpy mutant cells, it is likely that it diffuses away in the absence of the scaffold. Pio is not necessary to retain chitin at this stage, but it appears to have an analogous role later in development, when it is present in the pseudocone, to maintain the normal curved boundary of the inner surface of the corneal lens. Dpy and Pio have a similar function in the trachea, where they provide an elastic structural element of the lumen that controls its shape and size and maintains the chitin matrix. Removal of this structure by proteolysis is necessary for subsequent gas filling of the airway, and it is possible that the external corneal lens scaffold is likewise proteolytically degraded in late pupal stages. Transient aECM structures containing ZP-domain proteins also shape many cuticular and tubular structures in C. elegans, and α-tectorin on the surface of supporting cells templates the assembly of collagen fibrils in each layer of the tectorial membrane (Ghosh, 2024).
The arrangement of Dpy and Pio is dependent on Dyl; in wild-type ommatidia, they form linear structures, but this organization is not apparent in dyl mutants.The precise nature of these structures is not known, nor what additional components they may contain. ZP-domain proteins are capable of forming heteropolymeric filaments, and another ZP protein, Qsm, is known to promote the secretion and remodeling of Dpy filaments that link tendon cells to the pupal cuticle, increasing their tensile strength. Because Dyl is only present in the retina for a short time, it may be required to initiate the assembly of a scaffold structure that then becomes self-sustaining. Alternatively, because apical constriction is sufficient to alter Dpy organization and delay chitin accumulation, the role of Dyl may be limited to maintaining apical expansion. Nevertheless, the requirement for dyl for normal chitin deposition in bristles and wing hairs suggests that promoting chitin assembly, directly or indirectly, is one of its primary functions (Ghosh, 2024).
Other components of the corneal lens may also be affected by loss of dyl. Although chitin levels appear normal in sections of adult retinas, scanning electron micrographs show that the external surface of the corneal lens is incomplete. Induced apical constriction does not fully reproduce this loss of surface structure, suggesting that it reflects an independent function of Dyl. In the pupal wing, dyl belongs to a cluster of genes with peak expression at 42 hours APF, when the outer envelope layer of the cuticle is being deposited, but it influences the structure of inner layers as well. dyl expression in the embryo is also limited to a short time window in which apical cell remodeling and initial cuticle deposition occur (Fernandes, 2010). Another gene expressed at this time point, tyn, is required for the barrier function of the embryonic envelope layer. It is not clear whether the corneal lens has a typical cuticular envelope, but its outermost layer forms nanostructures composed of the Retinin protein and waxes. Although retinin mRNA is not strongly expressed until very late in pupal development, its expression is initiated at mid-pupal stages when Dyl is present. It is possible that Dyl and the Dpy-Pio scaffold also control the retention of Retinin and other components of the outer layer (Ghosh, 2024).
In summary, this study shows that the development of a biconvex corneal lens depends on both its upper and lower boundaries. The upper boundary, a convex shell of aECM that contains the ZP-domain proteins Dpy and Pio, encloses chitin and its associated proteins as they are secreted by cone and pigment cells. The shape of this shell may be defined by its attachment to the peripheral lattice cells and the pressure exerted on its center by secreted corneal lens components. The lower boundary is the apical plasma membrane of the cone and primary pigment cells. This membrane is flat until pseudocone secretion initiates late in pupal development, and the phenotype of dyl mutants suggests that it is maintained taut and expanded by attachment to the aECM. Loss of this expansion or of the pseudocone components Dpy or Pio gives the inner surface of the corneal lens a deeper curvature. It would be interesting to investigate whether mechanical forces exerted on the stromal ECM of the human cornea, perhaps through the ZP-domain proteins ZP4 and endoglin present in the underlying corneal endothelium, affect its shape and refractive power (Ghosh, 2024).
Biological systems are inherently noisy; however, they produce highly stereotyped tissue morphology. Drosophila pupal wings show a highly stereotypic folding through uniform expansion and subsequent buckling of wing epithelium within a surrounding cuticle sac (see Emergence of stereotypic folding within a cuticle sac). The folding pattern produced by buckling is generally stochastic; it is thus unclear how buckling leads to stereotypic tissue folding of the wings. This study found that the extracellular matrix (ECM) protein, Dumpy guides the position and direction of buckling-induced folds. Dumpy anchors the wing epithelium to the overlying cuticle at specific tissue positions. Tissue-wide alterations of Dumpy deposition and degradation yielded different buckling patterns. In summary, it is proposed that spatiotemporal ECM remodeling shapes stereotyped tissue folding through dynamic interactions between the epithelium and its external structures (Tsuboi, 2023).
This study has revealed that the spatiotemporally coordinated deposition and destruction of the ECM protein (Dpy) guide the position and direction of buckling. The results demonstrate that although cell populations show spatially homogeneous cellular behaviors (i.e., cell flattening), they can yield stereotypic tissue buckling morphology through the positional information encoded by ECM remodeling (Tsuboi, 2023).
Dpy, which anchors the wing tissue and cuticle, is constructed at specific positions (see Dumpy regulates the stereotypic buckling direction by connecting the wing epithelium and the pupal cuticle along veins). The position of Dpy anchorage is likely controlled posttranslationally, because Dpy overexpression did not result in the ectopic formation of Dpy anchorage. The construction of the Dpy matrix involves multiple processes, including synthesis, secretion, assembly into polymers, and association of the cuticle with the apical cell surface. Although the signaling pathway regulating Dpy anchoring remains unknown, the loss of vein Dpy in the mutant wing lacking veins suggests that positional information and signaling pathways related to wing vein patterning may contribute to the spatial differences in Dpy structure formation (Tsuboi, 2023).
Dpy degradation was found to initiate in the distal-posterior region, and the spatial pattern of degradation controls the position of the marginal fold (see Dumpy degradation is indispensable for the marginal fold formation). The degradation of Dpy is regulated by the action of Stubble (Sb) and Np apical transmembrane proteases, suggesting that the spatial pattern of Np/Sb expression or localization to the apical surface may control the degradation pattern of Dpy. Alternatively, another possible mechanism for the propagation pattern of Dpy degradation could be attributed to the spatial pattern of proteolytic activation of the proteases, because Np and Sb are expressed as catalytically inactive zymogens and require proteolytic cleavage to become active. In addition, regional differences in the physical properties of Dpy and its sensitivity to the proteases could contribute to the propagation pattern of Dpy degradation. It has been reported that Dpy undergoes filamentous conversion in response to increasing tension during indirect flight muscle development. Considering that Dpy filaments in wing tissue are formed under anisotropic proximal-distal–oriented tension caused by hinge contraction, the extent of Dpy filamentous conversion may vary regionally within the tissue depending on the applied stress. It would be interesting to investigate the regional difference in the physical properties of Dpy filaments within a tissue and their relevance to the propagation pattern of Dpy degradation (Tsuboi, 2023).
These findings are unique as they reveal the potential of external cues in generating stereotypic 3D tissue shapes within a spatially confined environment. This ECM-based mechanism should confer a potential means to generate diverse and controllable 3D tissue shaping in parallel with cell-intrinsic genetic programming. Future application of this ECM modification to tissue engineering would pave the way for manufacturing precisely folded tissues in any desired manner (Tsuboi, 2023).
Membrane expansion integrates multiple forces to mediate precise tube growth and network formation. Defects lead to deformations, as found in diseases such as polycystic kidney diseases, aortic aneurysms, stenosis, and tortuosity.
This study identified a mechanism of sensing and responding to the membrane-driven expansion of tracheal tubes. The apical membrane is anchored to the apical extracellular matrix (aECM) and causes expansion forces that elongate the tracheal tubes. The aECM provides a mechanical tension that balances the resulting expansion forces, with Dumpy being an elastic molecule that modulates the mechanical stress on the matrix during tracheal tube expansion. The zona pellucida (ZP) domain protein Piopio was shown to interact and cooperates with the ZP protein Dumpy at tracheal cells. To resist shear stresses which arise during tube expansion, Piopio undergoes ectodomain shedding by the Matriptase homolog Notopleural Tracheal tube lumen expansion requires mechanical stress regulation at apical cell membranes and attached aECM. This involves the proteolytic processing of proteins that set local membrane-matrix linkages. Thus, the membrane microenvironment exhibits critical roles in regulating tube and network functionality (Drees, 2023).
ZP domain proteins organize protective aECM in the kidney, tectorial inner ear, and ZP, as well as in Drosophila epidermis, tendon cells, and appendages. Drosophila ZP domain proteins link the aECM to actin and polarity complexes in epithelial cells. Dumpy establishes force-resistant filaments for anchoring tendon cells to the pupal cuticle (Drees, 2023).
This study found that ZP protein-mediated microenvironmental changes increase the flexibility of membrane-matrix association, resulting from the activity of ZP domain proteins. Shear stress stimulates the activity of membrane-anchored proteases and potentially also apical transmembrane protease Np since the misdistribution of the tracheal cytoskeleton was not observed when blisters arise. The dynamic membrane-matrix association control is based on findings that loss of Np prevents Pio ectodomain shedding at the apical cell membrane resulting in immobile localization of Pio at the membrane and Dpy localization within the matrix. Direct interaction and overlapping subcellular localization at the cell surface showed that both proteins form a ZP matrix that potentially attaches membrane and ECM. Deregulation of Pio shedding blocks ZP matrix rearrangement and release of membrane-matrix linkages under tube expansion and subsequent shear stress. This destabilizes the microenvironment of membranes, causing blister formation at the membrane due to ongoing membrane expansion. Additionally, Pio could be part of a force-sensing signal transduction system destabilizing the membrane and matrix. The observation that the membrane deformations are maintained in Np mutant embryos supports the postulated Np function to redistribute and deregulate membrane-matrix associations in stage 16 embryos when tracheal tube length expands. In contrast, Np overexpression potentially uncouples the Pio-Dpy ZP matrix membrane linkages resulting very likely in unbalanced forces causing sinusoidal tubes (Drees, 2023).
The membrane defects observed in both Pio and Np mutants indicate errors in the coupling of the membrane matrix due to the involvement of Pio. In pio mutants, gaps appear between the deformed membrane and the apical matrix. These changes in apical cell membrane shape are consistent with increased cell and tube elongation in pio mutant embryos because the matrix is uncoupled from the membrane in such mutants. In contrast to pio mutants, the large membrane bulges in Np mutants affect the membrane and the apical matrix. Since apical Pio is not cleaved in Np mutants, the matrix is not uncoupled from the membrane as in pio mutant embryos but is likely more intensely coupled, which leads to tearing of the matrix axially along the membrane bulges, when the tube expands in length. If apical Pio detachment reduces coupling between the matrix and apical membrane, then it is likely that Np mutant embryos may exhibit a reduced tube length phenotype. In Np mutant embryos, average tracheal dorsal trunk length tends to be reduced compared to wt embryos, suggesting that Pio shedding is critical in controlling tracheal tube lumen length (Drees, 2023).
Is Pio ectodomain shedding in response to tension? Tension was not measured directly. However, the developmental profile of mechanical tension during tracheal tube length elongation in stage 16 embryos is consistent with the profile of Pio shedding. Np cleaves apical Pio during stage 16 when tube length expands. In contrast, Pio shedding decreases sharply at early stage 17 when tube elongation is completed. The model, therefore, predicts that loss of Pio or increased Pio secretion at stage 16 may reduce the coupling of the membrane matrix so that increased tracheal tube elongation is maintained until the end of stage 16, which is found in pio mutants and upon Np overexpression. Unknown proteases may likely be involved in Pio processing since cleaved mCherry::Pio is also detectable in inactive NpS990A cells. Previously a mutation at the Pio ZP domain (R196A) resistant to NP cleavage in cell culture experiments (Drees, 2019). Establishing a corresponding mutant fly line would be essential in determining whether the observed phenotype resembles the phenotype of the Np mutant embryos. In addition, unknown mechanisms, such as distinct membrane connections during development and emerging links to the developing cuticle, may also influence tension at the apical membrane during tube length control (Drees, 2023).
Indeed, the anti-Pio antibody, which detects all different Pio variants, showed a punctuate Pio pattern overlapping with the apical cell membrane markers Crb and Uninflatable (Uif) at the dorsal trunk cells of stage 16 embryos. Additionally, Pio antibody also revealed early tracheal expression from embryonic stage 11 onward, and due to Pio function in narrow dorsal and ventral branches, strong luminal Pio antibody staining is detectable from early stage 14 until stage 17, when airway protein clearance removes luminal contents. In the pio5m and pio17c mutants, Pio stainings were strongly reduced although some puncta were still detectable in the trachea. Similarly, Pio antibody staining is intracellular in the trachea of stage 11 pio2R-16 point mutation embryos. Interestingly, also dpy mutants showed strongly reduced and intracellular Pio antibody staining (Drees, 2023).
mCherry::Pio was generated as a tool for in vivo Pio expression and localization pattern analysis during tube lumen length expansion. The mCherry::Pio resembled the Pio antibody expression pattern from early tracheal development onward. However, luminal mCherry::Pio enrichment occurs specifically during stage 16, when tubes expand. The stage 16 embryos showed mCherry::Pio puncta accumulating apically in dorsal trunk cells. Moreover, mCherry::Pio puncta partially overlapped with Dpy::YFP and chitin, a polymer of N-acetylglucosamine< at the taenidial folds, forming at apical cell membranes. Supported by several observations, such as antibody staining, video monitoring, FRAP experiments, and western blot studies, these findings indicate that Pio may play a significant role at the apical cell membrane and matrix in dorsal trunk cells of stage 16 embryo (Drees, 2023).
Furthermore, it was shown that Np mediates Pio ZP domain cleavage for luminal release of the short Pio variant during ongoing tube length expansion. The luminal cleaved mCherry::Pio is enriched at the end of stage 16 and finally internalized by the subsequent airway clearance process during stage 17 after tube length expansion. Such rapid luminal Pio internalization is consistent with a sharp pulse of endocytosis rapidly internalizing the luminal contents during stage 17. Wurst is required to mediate the internalization of proteins in the airways. Consistencly, during stage 17, luminal Pio antibody staining fades in control embryos but not in Wurst deficient embryos (Drees, 2023).
Nevertheless, Pio and its endocytosis depend on its interaction with the chitin matrix and the Np-mediated cleavage. In stage 16 wurst and mega mutant embryos, Pio antibody staining at the chitin cable, suggesting that Pio is cleaved and released into the dorsal trunk tube lumen. Also, the Cht2 overexpression did not prevent the luminal release of Pio. However, reduced wurst, mega function, and Cht2 overexpression caused an enrichment of punctuate Pio staining at the apical cell membrane and matrix . Although the three proteins are involved in different subcellular requirements, they all contribute to the determination of tube size by affecting either the apical cell membrane or the formation of a well-structured apical extracellular chitin matrix, indicating that changes at the apical cell membrane and matrix in stage 16 embryos affect the Pio pattern at the membrane. It also shows that local Pio linkages at the cell membrane and matrix are still cleaved by the Np function for luminal Pio release, which explains why those mutant embryos do not show pio mutant-like membrane deformations and Np-mutant-like bulges. This is in line with observations that tracheal Pio overexpression cannot cause tube size defects as the Np function is sufficient to organize local Pio linkages at the membrane and matrix. Therefore, it is unlikely that tracheal tube length defects in wurst and mega mutants as well as in Cht2 misexpression embryos are caused by the apical Pio density enrichment (Drees, 2023).
Nevertheless, oversized tube length due to the misregulation of the apical cell membrane and adjacent chitin matrix may cause changes to local Pio set linkages and the need for Np-mediated cleavage. Strikingly, a lack of Pio release was observed in Np mutants. This shows that Pio density at the membrane versus lumen depends predominantly on Np function. The molecular mechanisms that coordinate the Np-mediated Pio cleavage are unknown and will be necessary for understanding how tubes resist forces that impact cell membranes and matrices. On the other hand, Pio is required for the extracellular secretion of its interaction partner Dpy. At the same time, Dpy is needed for Pio localization at the cell membrane and its distribution into the tube lumen. Consistently, in vivo, mCherry::Pio, and Dpy::eYFP localization patterns overlap at the apical cell surface and within the tube lumen. These observations support the model that Pio and Dpy interact at the cell surface where Np mediates Pio cleavage to support luminal Pio release by the large and stretchable matrix protein Dpy (Drees, 2023).
Taenidial organization prevents the collapse of the tracheal tube. Therefore, cortical (apical) actin organizes into parallel-running bundles that proceed to the onset of cuticle secretion and correspond precisely to the cuticle’s taenidial folds. Mutant larvae of the F-actin nucleator formin DAAM show mosaic taenidial fold patterns, indicating a failure of alignment with each other and along the tracheal tubes. In contrast, pio mutant dorsal tracheal trunks contained increased ring spacing. Fusion cells are narrow doughnut-shaped cells where actin accumulates into a spotted pattern. Formins, such as Diaphanous, are essential in organizing the actin cytoskeleton. However, dorsal trunk tube fusion defects were not found as found in the presence of the activated Diaphanous (Drees, 2023).
On the other hand, ectopic expression of DAAM in fusion cells induces changes in apical actin organization but does not cause any phenotypic effects. DAAM is associated with the tyrosine kinase Src42A, which orients membrane growth in the axial tube dimension. Src42 overexpression elongates tracheal tubes due to flattened axially elongated dorsal trunk cells and AJ remodeling. Although flattened cells and tube overexpansion are similar in pio mutant embryos, mislocalization of AJ components was not observed, as found upon constitutive Src42 activation. Instead, an unusual stretched appearance of AJs was detected at the fusion cells of pio mutant dorsal trunks, which has not been observed before and may play a role in regulating axial taenidial fold spacing and tube elongation (Drees, 2023).
Self-organizing physical principles govern the regular spacing pattern of the tracheal taenidial folds. The actomyosin cortex and increased actin activity before and turnover at stage 16 drive the regular pattern formation. However, the cell cortex and actomyosin are in frictional contact with a rigid apical ECM. The Src42A mutant embryos contain shortened tube length but increased taenidial fold period pattern due to decreased friction. In contrast, the chitinase synthase mutant kkv1 has tube dilation defects and no regular but an aberrant pearling pattern caused by zero fiction (Drees, 2023).
In contrast, pio mutant embryos do not contain tube dilation defects or shortened tubes but increased tube length. Furthermore, chitin-binding probe (cbp) and antibody stainings reveal the presence of a luminal chitin cable and a solid aECM structure in pio mutant stage 16 embryos. In addition, apical actin enrichment in tracheal cells of pio mutant embryos appeared wt-like. Nonetheless, pio mutant embryos show an increased taenidial fold period compared with wt, indicating a decreased friction. Thus, it is proposed that the lack of Pio reduces friction. Reasons might be subtle defects of actomyosin constriction or chitin matrix, which was not detected in the pio mutant tracheal cells. Further reasons for lower friction might also be the loss of Pio set local linkages between apical cortex and aECM in stage 16 embryos, which are modified by Np, as proposed in in a model (see Model of apical Pio and Dpy matrix at the apical cell surface and Pio proteolysis and release. (Drees, 2023).
Heterozygous and homozygous pio mutant embryos generally do not show tubal collapse. However, the loss of Pio and accompanying lack of Dpy secretion in stage 17 pio mutant embryos led to the loss of a Pio/Dpy matrix, impacting the late embryonic maturation and differentiation of a normal chitin matrix at the apical cell surface. TEM images reveal reduced dense chitin matrix material at taenidial folds and misarranged taenidial fold pattern, suggesting impaired taenidial function prevents tube lumen from collapsing after tube protein clearance. wurst knockdown and mutant embryos do not show general tube collapse, but luminal chitin fiber organization is disturbed in stage 17 embryos. Therefore, transheterozygous wurst;pio mutant embryos may combine both defects and suffer from maturation deficits of the chitin/ZP matrix at the apical cell surface and within the tube lumen, which finally causes a high number of embryos with incomplete gas-filling due to tube collapse. These maturation deficits are even more dramatic in the wurst;pio double mutants, which show no gas-filling (Drees, 2023).
These studies on human Matriptase provide evidence for a mechanistic conservation of ZP domain protein as a substrate for ectodomain shedding. The upregulation of Matriptase activity and increased TGFβ receptor density affect human and mouse model idiopathic pulmonary fibrosis cells on pulmonary fibrogenesis. Furthermore, the human Matriptase induces the release of proinflammatory cytokines in endothelial cells, which contribute to atherosclerosis and probably also to abdominal aortic aneurysms. The membrane bulges arising in the Drosophila model during tracheal tube elongation upon Np loss of function showed analogy to the appearance of artery aneurysms. Bulges with varying phenotypic expression in different organs can lead to aortic rupture due to fragile artery walls or degeneration of layers in responses to stimuli, such as shear stresses. Indeed, aneurysm development is forced by alterations in the ECM and is characterized by extensive ECM fragmentation caused by shedding of membrane-bound proteins (Drees, 2023).
This study identified a dynamic control of matrix proteolysis, very likely enabling fast and site-specific uncoupling of membrane-matrix linkages when tubes expand. Such a scenario has not yet been studied in angiogenesis. It may represent a new starting point for genetic studies to decipher the putative roles of ZP domain proteins and Matriptase in clinically relevant syndromes, including the formation of aneurysms caused by membrane deformation and defects in size determination of airways and vessels (Drees, 2023).
Contractile tension is critical for musculoskeletal system development and maintenance. In insects, the muscular force is transmitted to the exoskeleton through the tendon cells and tendon apical extracellular matrix (ECM). In Drosophila, tendon cells were found to secrete Dumpy (Dpy), a zona pellucida domain (ZPD) protein, to form the force-resistant filaments in the exuvial space, anchoring the tendon cells to the pupal cuticle. Dpy undergoes filamentous conversion in response to the tension increment during indirect flight muscle development. Another ZPD protein Quasimodo (Qsm) was found to protect the notum epidermis from collapsing under the muscle tension by enhancing the tensile strength of Dpy filaments. Qsm is co-transported with Dpy in the intracellular vesicles and diffuses into the exuvial space after secretion. Tissue-specific qsm expression rescued the qsm mutant phenotypes in distant tissues, suggesting Qsm can function in a long-range, non-cell-autonomous manner. In the cell culture assay, Qsm interacts with Dpy-ZPD and promotes secretion and polymerization of Dpy-ZPD. The roles of Qsm underlies the positive feedback mechanism of force-dependent organization of Dpy filaments, providing new insights into apical ECM remodeling through the unconventional interaction of ZPD proteins (Chu, 2021).
At each stage of insect molt, epidermal morphogenesis takes place under the condition of their apical side being covered with apical ECM. Dpy is a major apical ECM component in the exuvial space required for proper morphogenesis of the adult notum, wing, male genitalia, and the embryonic trachea. Dpy distribution is non-uniform due to localized transcription and sometimes forms long strands connecting specific sites of the tissue and pupal cuticles. This study showed that Dpy forms filamentous structures that connect the apical surface of tendon cells to the cuticle. It is proposed that Dpy filaments support the notum epidermis from collapsing under the strong pulling force from the developing indirect flight muscles (Chu, 2021).
In the tendon structure of Dpy-YFP larvae, the YFP signal is arranged in a periodic pattern in parallel arrays of filaments, indicating that Dpy forms filaments by ordered association. The interval of YFP signal (0.55 μm) is significantly shorter than the length of Dpy polypeptide in a fully stretched form (3-8 μm, based on predicted Dpy length and the peptide bond length of 0.35 nm in beta-sheet configuration), suggesting that the Dpy protein is folded and assembled into a compact structure, and this may contribute to the tensile property of the filament. Furthermore, Dpy filaments are separated by a space of 0.67 μm that is filled with chitin. Those protein and polysaccharide complex would collectively stabilize the mature tendon matrix, the role previously ascribed to tonofilaments. However, the nearly uniform pattern of Dpy filaments filling the muscle insertion site appears different from the image of tonofilaments observed in late embryos that show a few distinct filaments under the transmission electron microscope.11 Whether Dpy is a structural component of tonofilaments remains to be evaluated (Chu, 2021).
Dpy matrix attached to tendon cells increased in amount and acquired filamentous organization under tension. The filamentous conversion of the Dpy matrix may involve tension-induced Dpy-matrix stretching and assembly of newly secreted Dpy into filaments. It is known that soluble Fibronectin (FN) dimers are converted into fibril under cytoskeletal tension applied to the FN receptor integrin; the tension-dependent FN remodeling phenomenon was also observed in Xenopus and zebrafish tissues (Chu, 2021).
We identified Qsm as a factor required for Dpy remodeling. In normal qsm+ pupae, Dpy-YFP filaments spanning the exuvial space opened up to ∼30 μm. Without qsm, Dpy filaments were overstretched at the high tension status (32∼34 h APF), opening the exuvial space up to 3-fold. Therefore, it is proposed that muscle-derived tension and Qsm-dependent modulation are the major sources of mechanical and chemical effectors of Dpy matrix maturation, respectively (Chu, 2021).
Given the critical requirement for Dpy matrix rigidity and the presence of ZPD, a signature of structural protein, the diffusible nature of Qsm in the exuvial space was unexpected. The potential action of Qsm in the protein secretory pathway of Dpy is discussed (Chu, 2021).
To study the properties of ZPDs, a mini-construct mVenus-Dpy-ZPD was used, which was cleaved to release ZPD that was secreted and formed oligomers in cell culture. The association of Qsm with the membrane-bound form of Dpy-ZPD inside the cell was consistent with the observation of co-transportation of Dpy and Qsm in the tendon cells. In contrast, only a minimal level of complex formation by extracellular Dpy-ZPD and Qsm was detected. These data suggest that the interaction between Dpy-ZPD and Qsm occurs as immature membrane-bound forms prior to the formation of proper disulfide bonds. The complex of Dpy-ZPD and Qsmmut may stall as maturation intermediates and are degraded as misfolded proteins. In this respect, Qsm functions as a specific co-factor, a molecular chaperon, and is likely important for the conformational maturation of Dpy-ZPD (Chu, 2021).
A possible mechanism is proposed based on an in vitro assay. After Qsm and Dpy-ZPD acquire the proper conformation and disulfide bonds, they dissociate from each other, separate from the membrane following the Furin cleavage, and are delivered to the protein secretory pathway. Dissociation of the Dpy-Qsm complex after this stage is probably important for Dpy maturation since the excessive expression of Qsm was inhibitory to the oligomerization of Dpy-ZPD. As the majority of Dpy consists of a large number of EGF repeats, Dpy maturation and secretion may involve additional mechanisms. Proper deposition and maturation of this gigantic protein is complex, involving regulation at the ER exit site by Tango1, glycosylation and perhaps many other regulators. Another example of ZPD proteins that require each other for their normal secretion is NOAH-1 and NOAH-2 in C. elegans, which are the apical ECM proteins required for epidermal integrity and muscle anchoring (Chu, 2021).
In qsm mutants, expressed Qsm rescued defects in distant locations, suggesting that secreted Qsm diffuses in the exuvial space and interacts with the target tissues to support proper Dpy filament formation. Tendon cells were observed to incorporate mCherry-Qsm expressed in an ectopic tissue (muscle) and package into cellular vesicles containing Dpy-YFP that are transported to the interface of tendon cells and apical-ECM. This delivery process may involve endocytosis and retrograde recycling through the ER-Golgi compartment, as previously shown for the secreted enzyme chitin deacetylase Serpentine in the remodeling of the tracheal apical ECM. Dpy-YFP co-secreted with mCherry-Qsm may be incorporated into Dpy filaments by the secretion and capture mechanism. Diffusing Qsm would be incorporated by distant tissues and regulate the Dpy maturation and secretion. In addition, diffused Qsm may directly interact with extracellular Dpy to promote the remodeling of the Dpy matrix, although extracellular Dpy-Qsm interaction remains to be established. Fusion of mCherry somewhat compromised the long-range rescue activity of Qsm. This may be due to the reduction of the diffusion constant, or the mCherry moiety interfering with retrograde transport (Chu, 2021).
qsm was first identified based on the screening for circadian regulated genes (Stempfl, 2002). Subsequent studies revealed that qsm is expressed in clock neurons in the adult brain in a light-inducible manner. It was hypothesized that Qsm works cell-autonomously to regulate the degradation of the transcription factor Timeless (Chen, 2011). However, how this model fits the finding of Qsm as a soluble extracellular protein and a modulator of Dpy is unknown. Thus, further studies on the behavior of Qsm protein in the extracellular space and intracellular compartment, and additional interaction targets of Qsm will be required to understand the full picture of Qsm function (Chu, 2021).
In conclusion, this study identified Qsm as a novel class of ZPD proteins that acts non-structurally to support the apical ECM maturation under mechanical force. Diffusion of Qsm allows whole-body modification of polymer formation of Dpy and possibly other ZPD proteins. Since the timing of Qsm expression during cuticle formation is tightly regulated under the ecdysis program, Qsm may be an excellent candidate for a systemic modulator of apical ECM at the time of dynamic tissue movement of metamorphosis. Recent studies have uncovered ECM modulation mechanisms by regulated synthesis and secretion, subunit assembly, chemical modification, and proteolytic degradation. Qsm is a new class of molecules regulating apical ECM dynamics by targeting a subset of ZPD proteins and is expected to provide a new direction in the developmental control of ECM dynamics (Chu, 2021).
The Drosophila extracellular matrix protein Dumpy (Dpy) is one of the largest proteins encoded by any animal. One class of dpy mutations produces a characteristic shortening of the wing blade known as oblique (dpyo), due to altered tension in the developing wing. This study describes the characterization of docked (doc), a gene originally named because of an allele producing a truncated wing. This study shows that doc corresponds to the gene model CG5484, which encodes a homolog of the yeast protein Yif1 and plays a key role in ER to Golgi vesicle transport. Genetic analysis is consistent with a similar role for Doc in vesicle trafficking: docked alleles interact not only with genes encoding the COPII core proteins Sec23 and Sec13, but also with the SNARE proteins Synaptobrevin and Syntaxin. Further, it was demonstrated that the strong similarity between the doc1 and dpy0 wing phenotypes reflects a functional connection between the two genes; various dpy alleles were found to be sensitive to changes in dosage of genes encoding other vesicle transport components such as Sec13 and Sar1. Doc's effects on trafficking are not limited to Dpy; for example, reduced doc dosage disturbed Notch pathway signaling during wing blade and vein development. These results suggest a model in which the oblique wing phenotype in doc1 results from reduced transport of wild-type Dumpy protein; by extension, an additional implication is that the dpy0 alleles can themselves be explained as hypomorphs (Kandasamy, 2021).
One of the fundamental gaps in knowledge of how novel anatomical structures evolve is understanding the origins of the morphogenetic processes that form these features. This study traced the cellular development of a recently evolved morphological novelty, the posterior lobe of D. melanogaster (see The posterior lobe protrudes from the lateral plate.). This genital outgrowth forms through extreme increases in epithelial cell height. By examining the apical extracellular matrix (aECM), a vast matrix was found associated with the developing genitalia of lobed and non-lobed species. Expression of the aECM protein Dumpy is spatially expanded in lobe-forming species, connecting the posterior lobe to the ancestrally derived aECM network. Further analysis demonstrated that Dumpy attachments are necessary for cell height increases during posterior lobe development. It is proposed that the aECM presents a rich reservoir for generating morphological novelty and highlights a yet unseen role for aECM in regulating extreme cell height (Smith, 2020).
This work demonstrates an important role for the aECM protein Dumpy in the development of the posterior lobe, as exhibited by the dramatic phenotypes in the dumpy RNAi background and the strong association of Dumpy:YFP with only the tallest cells in these experiments. The presence of Dumpy:YFP at the ventral tip of the defective posterior lobe in dumpy RNAi individuals indicated that the driver does not supply sufficient RNAi in ventral tip cells to remove all of the Dumpy-YFP, though a strong reduction in florescence is observed at the ventral tip. This ventral tip would not fully form if Dumpy is completely removed, however, there is no driver that can test this prediction (Smith, 2020).
Overall, the data are consistent with three possible mechanisms that could allow Dumpy to regulate cell height. First, Dumpy could serve as a structural support while autonomous cell mechanical processes drive apico-basal elongation, such as by altering the cytoskeleton, which is observed in posterior lobe cells. Second, the cells of the posterior lobe could be pulled mechanically through their connection to the Dumpy aECM. This process could operate passively, deforming cells of the lobe, but could also drive changes in the cytoskeleton in response to external tensions. Finally, the aECM could alter cell signaling dynamics, as has been exhibited by the basal ECM. Previous research has shown that the JAK/STAT pathway is important for posterior lobe development, and the ability for a signal to reach its target cells could be altered in the absence of Dumpy. Of course, these models are not mutually exclusive and some combination of these mechanisms may be integrated to shape the posterior lobe. Observations of increased cytoskeletal components in posterior lobe cells and the reduced height of cells that lack Dumpy in knockdown experiments are consistent with all three mechanisms, which are difficult to differentiate experimentally (Smith, 2020).
Genitalia represents some of the most rapidly diversifying structures in the animal kingdom, and the results suggest the aECM may participate in the modification of Drosophila genital structures. The shape of the posterior lobe is extremely diverse among species of the melanogaster clade. It was confirmed that the extended posterior lobe of D. sechellia is associated with a similar aECM to D. melanogaster, confirming that this mechanism is likely common to other lobed species. The results demonstrate that reducing the levels of Dumpy can affect the shape of the posterior lobe, with extreme knockdown phenotypes approximating the posterior lobe of D. mauritiana. Furthermore, the sheath and phallus show dense deposits of Dumpy, suggesting that the aECM could play important roles in diversifying these remarkably variable structures. During the course of evolution, one could imagine that by altering which cells are connected to the aECM, the physical nature of those connections, and the mechanical properties of those cells and/or their associated ECM could lead to changes in morphological shape. Hence, identifying genes that have evolved to differentiate these structures could uncover novel mechanisms for genetically controlling the behavior of this aECM and behaviors of cells bound to this dynamic scaffold (Smith, 2020).
Evolution is thought to act through the path of least resistance. When confronted with the remarkable diversity of genital morphologies present in insects, one must wonder how the intricate projections, bumps, and divots form through the action of epithelial rudiments. Traditionally the field of evo-devo research has focused on cell intrinsic processes that contribute to tissue morphogenesis, but this study shows that the extrinsic forces from the aECM may be important for the evolution of a novelty. The aECM, while understudied, has been implicated in the morphogenesis of many structures, making it a promising target for evolution. A conserved aECM network associated with central genital structures (clasper, sheath, and phallus) is found in both lobed and non-lobed species. However, on the dorsal side of the lateral plate differences are observed, with lobed species having substantial deposits of aECM that fill this area between the lateral plate and clasper and non-lobed species forming only thin strand-like connections. Because a complex network of aECM already existed on the genitalia to potentially pattern other structures, such as the phallus and its multiple elaborations, it is hypothesized that this spatial expansion of aECM to the posterior lobe cells allowed them to be integrated into the ancestral aECM network, a step which was likely significant to its evolution. Overall, this suggests that the aECM could be an unexpected target for generating novel anatomical structures. On the other hand, tissues which lack such an ancestral aECM network may be less likely to evolve projections through this mechanism. In addition, while Dumpy may be required for the development of the posterior lobe, additional components of the aECM, including factors that remodel the aECM or receptors that anchor the aECM to the cells, are likely also needed. Furthermore, it is envisioned that additional processes may also be contributing to the full morphogenesis of the posterior lobe, including potential intrinsic processes that may contribute to the cell shape changes that are observed, such as the elevated concentrations of cytoskeletal components (Smith, 2020).
The expanded dumpy expression discovered in this study have led to a consideration of how the posterior lobe gained this aECM attachment. Models of gene network co-option have been appealing because they establish pre-existing mechanisms in place that can be rapidly ported to new locations to generate massive changes in a tissue. Interestingly, previous work uncovered a gene regulatory network (GRN) that regulates development of an ancestral embryonic structure, the posterior spiracles, which was co-opted during the evolution of the posterior lobe and regulates its development (Glassford, 2015). Along these lines, dumpy is expressed in the developing posterior spiracles (Wilkin, 2000), and a thin strand of Dumpy:YFP was observed connecting the posterior spiracles to the surrounding embryonic cuticle, known as the vitelline membrane. Identification of regulatory elements which activate dumpy in the posterior lobe will be necessary to determine whether its role in the posterior spiracle was relevant to the evolution of expanded genital expression (Smith, 2020).
Identifying the genetic changes that regulate cellular processes required for the evolution of new structures represents a major looming challenge in evo-devo research. Taken as a whole, the posterior lobe offers a promising model to begin to understand how gene regulatory networks and terminal effectors are connected, in addition to understanding how these networks become associated with novelties during evolution. As understanding of the patterning of these tissues both at the level of transcriptional regulators and terminal effectors (such as Dumpy) becomes more complete, it is possible to begin to connect the regulation of gene expression with the developmental behaviors of different cell types. The mapping of regulatory enhancers through reporter assays will allow tracing of the evolutionary history of these connections (Glassford, 2015). Furthermore, in systems where sophisticated genetic modifications are possible, CRISPR/Cas9 can be used to test the phenotypic consequences of genetic variation in these regulatory enhancers by exchanging regions between species. Coupling such manipulations to quantitative measures of morphogenesis will illuminate the root causes of differences in tissue architecture (Smith, 2020).
Extracellular matrix (ECM) assembly and remodelling is critical during development and organ morphogenesis. Dysregulation of ECM is implicated in many pathogenic conditions, including cancer. The type II transmembrane serine protease matriptase and the serine protease prostasin are key factors in a proteolytic cascade that regulates epithelial ECM differentiation during development in vertebrates. This study shows by rescue experiments that the Drosophila proteases Notopleural (Np) and Tracheal-prostasin (Tpr) are functional homologues of matriptase and prostasin, respectively. Np mediates morphogenesis and remodelling of apical ECM during tracheal system development and is essential for maintenance of the transepithelial barrier function. Both Np and Tpr degrade the zona pellucida-domain (ZP-domain) protein Dumpy, a component of the transient tracheal apical ECM. Tpr zymogen and the ZP domain of the ECM protein Piopio are cleaved by Np and matriptase in vitro. These data indicate that the evolutionarily conserved ZP domain, present in many ECM proteins of vertebrates and invertebrates, is a novel target of the conserved matriptase-prostasin proteolytic cascade (Drees, 2019).
This paper reports that the vertebrate matriptase-prostasin proteolytic cascade, which is crucial for extracellular matrix differentiation and tissue homeostasis, is conserved in Drosophila. Np acts as a functional Drosophila homologue of matriptase, and tpr mediates prostasin function in the Drosophila tracheal system. Cleavage targets of these conserved extracellular proteolytic pathways are the ZP domains, present in many extracellular proteins of both vertebrates and invertebrates. The Np-mediated protease cascade controls at least three distinct cellular processes during tracheal development, i.e. morphogenesis of the taenidial folds, degradation of the tracheal Dpy cable in the tracheal lumen, and maintenance of the transepithelial barrier function (Drees, 2019).
In vertebrates, proteolysis by matriptase plays a key role in regulating epithelial differentiation. Ectopic expression of human matriptase in the developing tracheal system of Np mutant embryos rescues all aspects of the Np mutant phenotype, i.e. degradation of the Dpy luminal cable, taenidial folds formation, and gas filling of the tubes. The fact that matriptase can functionally substitute for the lack of Np activity indicates that the two proteins share similar functions. Furthermore, similar to matriptase, Np is differentially localized in tissue and stage-dependent patterns at the apical plasma membrane and the apical extracellular space. These findings suggest processing of Np by ectodomain shedding similar to what has been described for matriptase. However, the processing of human matriptase involves not only ectodomain shedding but also the transient interaction of the stem region with its cognate inhibitor HAI-2. Absence of HAI-2 prevents cell surface translocation of matriptase and causes its accumulation in the Golgi compartment (Lai, 2015). Ectopic HAI-2 expression in combination with matriptase facilitates secretion of matriptase in the Drosophila tracheal system. This finding and the lack of a Drosophila HAI-2 homologue indicate different regulatory mechanisms for the translocation and ectodomain shedding of Np and matriptase. This assumption is also consistent with the lack of conserved regulatory stem regions of Np and matriptase. Therefore, different protein processing mechanisms of the two otherwise functionally equivalent proteins are proposed. However, the apparent diverse regulation of both proteins provides the possibility to establish an in vivo experimental system to analyse aspects of matriptase regulation and processing in Drosophila (Drees, 2019).
The matriptase-prostasin proteolytic cascade is initiated by rapid matriptase autoactivation as shown by an in vitro cell-free system as was observed with Np. Thus, the proteolytic cascade involved in aECM formation and maturation in Drosophila is likely to be initiated by Np autoactivation. Once activated, it acts on Tpr, a direct downstream target of Np in the developing tracheal system. This conclusion is based on the observation that in vitro-purified Np is able to cleave Tpr at the zymogen activation site, implying a direct activation of Tpr zymogen by Np in vivo. Furthermore, the tracheal aECM phenotype of tpr mutant embryos is less pronounced than the Np phenotype, since taenidial folds are wild-type like in tpr mutant embryos, while Np mutant embryos lack taenidial folds. The observation that human matriptase also cleaves Tpr at the same zymogen activation site provides additional support for the functional identity of matriptase and Np (Drees, 2019).
Ectopic tracheal expression of human prostasin, together with human HAI-2 in tpr mutant embryos, rescues the defects of the aECM and the LC phenotype of tpr mutants. Thus, Tpr is a functional homologue of human prostasin in the developing trachea. Also, human HAI-2 is required for prostasin secretion into the tracheal lumen, as has been reported for prostasin in vertebrate tissues [43]. LC defects, as observed in tpr mutants, are often caused by an impaired transepithelial barrier function. However, tpr mutant embryos develop a normal barrier and, thus, it is supposed that the LC defects are likely caused by hampered degradation of luminal material and/or Tpr might affect epithelial sodium channels (ENaCs). ENaCs are located in the apical membrane of tracheal cells and are critical for tracheal gas fillin. In vertebrates, prostasin activates ENaCs by inducing proteolytic cleavage of the gamma subunit. It will be interesting to see whether Tpr plays a similar role for ENaC activation in Drosophila (Drees, 2019).
Vertebrate prostasin is widely expressed in ectodermal tissue and most functional aspects of human matriptase are mediated via prostasin in the various tissues. However, the functional relationship of matriptase and prostasin remains to be clarified since matriptase activation and shedding is prostasin-dependent in specific tissues. In contrast to vertebrate prostasin, Drosophila Tpr expression is confined to the tracheal system and not detectable in other ectodermal tissues. Thus, while vertebrate matriptase and prostasin are co-expressed in most tissues, a different scenario is proposed in Drosophila. Tpr belongs to a small group of seven very similar proteases in Drosophila. The corresponding genes are differentially expressed in specific spatial patterns in various ectodermal tissues.It is proposed that such protease zymogens represent putative Np cleavage targets that mediate Tpr-like functions in the different ectodermal tissues that express Np including salivary glands, hindgut, and epidermis (Drees, 2019).
Drosophila tracheal development is a paradigm for the generation of branched tubular systems. Early steps of tracheal maturation, notably tube formation and tubular network assembly, develop independently of Np, while tracheal aECM formation and the transepithelial barrier function during late embryonic tracheogenesis depend on Np. Main differentiation events of the aECM, such as taenidial folds morphogenesis and degradation of luminal protein matrix, are controlled by Np and Tpr (Drees, 2019).
The taenidial folds of the tracheal aECM mainly consist of chitin running perpendicular to the tracheal tube length along the lumen. Their main function is to provide stiffness combined with concurrent flexibility of the tube. Taenidial folds formation is severely affected in Np mutant embryos. The outmost taenidial structure, the hydrophobic envelope, is not detectable and the chitin strands of taenidial folds are highly disorganized. Chitin-interacting proteins, involved in chitin organization, may represent putative targets of Np activity. The aECM phenotype of tpr mutant embryos suggests that tpr function is more specific and confined to establish a proper adhesion between the apical side of tracheal cells and the overlaying taenidial folds of the aECM (Drees, 2019).
Both Np and Tpr are also involved in the degradation of tracheal luminal Dpy, a large ZP domain-containing protein. Luminal Dpy is part of a chitin-proteinous matrix within the tracheal lumen and is essential for normal tracheal network and tube formation. The luminal matrix is degraded and removed from the tracheal lumen during formation of the tracheal taenidial folds and the subsequent gas filling of the tracheal system. Np appears to be the key factor in Dpy cable degradation since Np-deficient embryos completely lack Dpy degradation. Some degradation, however, is mediated via Tpr proteolysis because tpr mutants display remaining Dpy material in the tracheal lumen during late embryogenesis. Thus, the combination of Np and Tpr accomplish luminal Dpy degradation. Alternatively or in addition, Np activates unknown proteases that mediate complete Dpy degradation prior to the gas filling of the tracheal tubes (Drees, 2019).
The transepithelial barrier function is established by the septate junction (SJ) protein complexes, localized at the apico-lateral membrane of epithelial cells. The lack of bona fide SJ proteins like the Drosophila claudin Mega causes a disruption of the ladder-like SJ ultrastructure and a barrier function defect. In Np mutants, the ladder-like ultrastructure of SJs and the barrier function appear to be properly established, but the barrier function collapses during the end of embryogenesis. Thus, Np is essential for the maintenance of the transepithelial barrier function mediated by SJs. This function of Np is reminiscent of matriptase function in mammals. Tracer injection experiments into the dermis of matriptase-deficient mice indicate impaired epidermal tight junction function in such animals. In intestinal epithelial model cell layers and hypomorphic matriptase mice, the essential tight junction component Claudin-2 is deregulated. This observation suggests that reduced barrier integrity was caused, at least in part, by an impaired claudin-2 protein turnover [51]. Furthermore, matriptase cleaves EpCAM, which in turn decreases EpCAM ability to associate with claudin-7 followed by lysosomal degradation of claudin-7. Thus, it is speculated that Np may also control the maintenance of the epithelial barrier in Drosophila by regulating the function or turnover of claudins in SJs, the invertebrate analogue of the vertebrate tight junction (Drees, 2019).
The aECM protein Dpy is an in vivo downstream target of both Np and human matriptase proteolytic activity. This observation was puzzling since Dpy is not conserved in vertebrates. However, Dpy contains a conserved region, the ZP domain. The ZP domain defines a conserved family of aECM proteins, originally identified in the zona pellucida coat surrounding the mammalian oocyte. The 260 amino acids long ZP domain is proposed to act as a module promoting polymerization of proteins into threads and matrices essential for the organization of highly specialized apical extracellular structures. In fact, the ZP domain was confirmed as a target of Np and matriptase by data showing that Pio, an aECM protein containing a ZP domain, is cleaved by Np and matriptase. Both cleave the ZP domain of Pio within the short linker region, which separates ZP-N and ZP-C, the two subdomains of the ZP domain. Pio is secreted apically in the tracheal lumen and establishes together with Dpy, possibly via ZP-domain polymerization, a structural matrix in the tracheal lumen that is essential for the formation of an interconnected branched network. Cleavage of ZP domains within a meshwork of Dpy and Pio filaments may facilitate rapid degradation of the luminal extracellular matrix, the prerequisite for normal gas filling of the tracheal system. This conclusion is supported by phenotypic analysis of Np mutant embryos, which exhibit a stable, undegraded luminal Dpy cable and lack tracheal gas filling. ZP-domain proteins also play crucial roles in development of embryonic epidermal cuticle, an aECM that protects the animal against the external milieu. Eight ZP-domain proteins are required for the localized reorganization of epidermal cells and to sculpture the actin-rich apical extensions, the denticles. The observation that Np mutants exhibit reduced and rudimentary denticles in the epidermis underlines the possibility that the epidermal ZP-proteins are also targets of Np protease activity (Drees, 2019).
The results showing that human matriptase cleaves the ZP domain of Pio open future directions to explore novel targets of the matriptase-prostasin catalytic pathway. In vertebrates, ZP-domain proteins are involved in remodelling apical extracellular structures, such as ZP1-ZP3, important in the mammalian ovary for fertilization and uromodulin, which is released into the tubular kidney lumen where it polymerizes in a gel-like matrix that controls salt transport and urine concentration. Also, mutations in genes encoding ZP-domain proteins cause human diseases such as deafness, triggered by mutations in alpha- and beta-tectorin. The tectorins are components of the tectorial membrane, an aECM necessary for sound transmission to neural cells in the cochlea. These examples already demonstrate the importance of ZP-domain proteins for mammalian physiology. Based on the results reported in this study, it is proposed that ZP-domain cleavage by the matriptase-prostasin proteolytic cascade may represent a conserved process to control ZP-domain protein functions, which are crucial for apical matrix remodelling during development, wound repair, and differentiation (Drees, 2019).
Tango1 enables ER-to-Golgi trafficking of large proteins. Loss of Tango1, in addition to disrupting protein secretion and ER/Golgi morphology, causes ER stress and defects in cell shape. The previously observed dependence of smaller cargos on Tango1 is a secondary effect. If large cargos like Dumpy, which this study identifies as a Tango1 cargo, are removed from the cell, nonbulky proteins reenter the secretory pathway. Removal of blocking cargo also restores cell morphology and attenuates the ER-stress response. Thus, failures in the secretion of nonbulky proteins, ER stress, and defective cell morphology are secondary consequences of bulky cargo retention. By contrast, ER/Golgi defects in Tango1-depleted cells persist in the absence of bulky cargo, showing that they are due to a secretion-independent function of Tango1. Therefore, maintenance of ER/Golgi architecture and bulky cargo transport are the primary functions for Tango1 (Rios-Barrera, 2017).
The endoplasmic reticulum (ER) serves as a major factory for protein and lipid synthesis. Proteins and lipoproteins produced in the ER are packed into COPII-coated vesicles, which bud off at ER exit sites (ERES) and then move toward the Golgi complex where they are sorted to their final destinations. Regular COPII vesicles are 60-90 nm in size, which is sufficient to contain most membrane and secreted molecules. The loading of larger cargo requires specialized machinery that allows the formation of bigger vesicles to accommodate these bulky molecules. Tango1 (Transport and Golgi organization 1), a member of the MIA/cTAGE (melanoma inhibitory activity/cutaneous T cell lymphoma-associated antigen) family, is a key component in the loading of such large molecules into COPII-coated vesicles. Molecules like collagens and ApoB (apolipoprotein B)-containing chylomicrons are 250-450 nm long and rely on Tango1 for their transport out of the ER, by physically interacting with Tango1 or Tango1 mediators at the ERES (Rios-Barrera, 2017).
Tango1 is an ER transmembrane protein that orchestrates the loading of its cargo into vesicles by interacting with it in the ER lumen. The interaction of Tango1 with its cargo then promotes the recruitment of Sec23 and Sec24 coatomers on the cytoplasmic side, while it slows the binding of the outer layer coat proteins Sec13 and Sec31 to the budding vesicle. This delays the budding of the COPII carrier. Tango1 also recruits additional membrane material to the ERES from the Golgi intermediate compartment (ERGIC) pool, thereby allowing vesicles to grow larger. It also interacts directly with Sec16, which is proposed to enhance cargo secretion. A shorter isoform of mammalian Tango1 lacks the cargo recognition domain but nevertheless facilitates the formation of megacarrier vesicles (Rios-Barrera, 2017).
Apart from bulky proteins, some heterologous, smaller proteins like secreted horseradish peroxidase (ssHRP, 44 kDa) and secreted GFP (27 kDa) also depend on Tango1 for their secretion . Unlike for collagen or ApoB, there is no evidence for a direct interaction between Tango1 and ssHRP or secreted GFP. It is not clear why Tango1 would regulate the secretion of these molecules, but it has been proposed that in the absence of Tango1, the accumulation of nonbulky proteins at the ER might be due to abnormally accumulated Tango1 cargo blocking the ER; however, this has not been tested experimentally (Rios-Barrera, 2017).
Drosophila Tango1 is the only member of the MIA/cTAGE family found in the fruit fly, which simplifies functional studies. Like vertebrate Tango1, the Drosophila protein participates in the secretion of collagen. And as in vertebrates, ssHRP, secreted GFP, and other nonbulky molecules like Hedgehog-GFP also accumulate in the absence of Tango1. These results have led to the proposal that Tango1 participates in general secretion. However, most of the evidence for these conclusions comes from overexpression and heterologous systems that might not reflect the physiological situation (Rios-Barrera, 2017).
This study describes a tango1 mutant allele that was identified in a mutagenesis screen for genes affecting the structure and shape of terminal cells of the Drosophila tracheal system. Tracheal terminal cells form highly ramified structures with branches of more than 100 μm in length that transport oxygen through subcellular tubes formed by the apical plasma membrane. Their growth relies heavily on membrane and protein trafficking, making them a very suitable model to study subcellular transport. Terminal cells were used to study the function of Tango1, and loss of Tango1 was found to affect general protein secretion indirectly, and it also leads to defects in cell morphology and in the structure of the ER and Golgi. The defects in ER and Golgi organization of cells lacking Tango1 persist even in the absence of Tango1 cargo (Rios-Barrera, 2017).
These studies led to an explanation of why, in the absence of Tango1, nonbulky proteins accumulate in the ER despite not being direct Tango1 cargos. These cargos are retained in the ER as a consequence of nonsecreted bulky proteins interfering with their transport. However, the effect of loss of Tango1 on ER/Golgi morphology can be uncoupled from its role in bulky cargo secretion (Rios-Barrera, 2017).
This study has described a role of Tango1, which was initially identified through its function in tracheal terminal cells and other tissues in Drosophila embryos, larvae, and pupae. Due to their complex shapes and great size, terminal cells are a well-suited system to study polarized membrane and protein trafficking, with the easily scorable changes in branch number and maturation status providing a useful quantitative readout that serves as a proxy for functional membrane and protein trafficking machinery. Moreover, this analyses are conducted in the physiological context of different tissues in the intact organism (Rios-Barrera, 2017).
The loss-of-function allele tango12L3443 has a stop codon eight amino acids downstream of the PRD domain and eliminates the 89 C-terminal amino acids of the full-length protein. It is unlikely that the mutation leads to a complete loss of function. First, terminal cells expressing an RNAi construct against tango1 show stronger defects, with fewer branches per cell than homozygous tango12L3443 cells. Second, the mutant protein appears not to be destabilized nor degraded, but instead is present at apparently normal levels, albeit at inappropriate sites. Predictions of the deleted fragment of the protein suggest it is disorganized and that it contains an arginine-rich domain that has no known interaction partners and that is not present in human Tango1. In homozygous mutant terminal cells, the mutant tango12L3443 protein fails to localize at ERES. In mammalian Tango1, the Sec16-interacting region within the PRD domain is necessary for the localization of Tango1 to the ERES and for its interaction with Sec23 and Sec16, but since this domain is fully present in tango12L3443, the results mean that either the missing 89 C-terminal amino acids contain additional essential localization signals, or that the PRD domain is structurally affected by the truncation of the protein. The latter is considered less likely, as a truncation of eight amino acids downstream of the PRD domain is unlikely to destabilize the polyproline motifs, especially as the overall stability of the protein does not seem to be affected. Furthermore, this region shows a high density of phosphoserines (Ser-1345, Ser-1348, Ser-1390, and Ser-1392), suggesting it might serve as a docking site for adapter proteins or other interactors (Rios-Barrera, 2017).
Terminal cells lacking Tango1 have fewer branches than control cells and are often not properly filled with air. This loss-of-function phenotype is not due to a direct requirement for Tango1, as it is suppressed by the simultaneous removal of Dumpy (Dpy), an extracellular protein involved in epidermal-cuticle attachment, aposition of wing surfaces and trachea development. It also cannot be explained by the individual loss of crb, Piopio (Pio) a zona pellucida (ZP) domain protein that mediates the adhesion of the apical epithelial surface and the overlying apical extracellular matrix, or dpy, since knocking down any of these genes has no effect on cell morphology. Instead, it is proposed that the cell morphological defect is due at least in part to the activation of the ER stress response, since expression of Xbp1 is sufficient to recapitulate the phenotype. Xbp1 regulates the expression of genes involved in protein folding, glycosylation, trafficking, and lipid metabolism. It is possible that one or a small number of specific genes downstream of Xbp1 are responsible for defective branch formation or stability, but the phenotype could also be a secondary consequence of the physiological effects of the ER stress response itself, for example, a failure to deliver sufficient lipids and membrane from the ER to the apical plasma membrane (Rios-Barrera, 2017).
Collagen, with a length of 300 nm, and ApoB chylomicrons with a diameter of > 250 nm, have both been biochemically validated as Tango1 cargos. These molecules are not expressed in terminal cells, and therefore it was clear that Tango1 must have a different substrate in these cells. Given that Tango1 is known for the transport of bulky cargo, that Dpy is the largest Drosophila protein at 800 nm length, and that Dpy vesicles are associated with Tango1 rings in tracheal cells, it is proposed that Dpy is a further direct target of Tango1. Colocalization of Tango1 with its cargo has also been observed in other tissues: with collagen in Drosophila follicle cells and with ApoB in mammalian cell lines (Rios-Barrera, 2017).
No regions of sequence similarity that could represent Tango1-binding sites have been found in Tango1 cargos. There are several possible explanations for this. First, these proteins may contain binding motifs, but the motifs are purely conformational and not represented in a linear amino acid sequence. There is no evidence for or against this hypothesis, but it would be highly unusual, and there is support for alternative explanations. Thus, as a second possibility, all three proteins may require Tango1 for their secretion, but variable adapters could mediate the interactions. In vertebrates, Tango1 can indeed interact with its cargo through other molecules; for instance, its interaction with collagen is mediated by Hsp47. However, in Drosophila, there is no Hsp47 homolog. In the case of ApoB, it has been suggested that microsomal triglyceride transfer protein (MTP) and its binding partner, protein disulphide isomerase (PDI), might associate with Tango1 and TALI to promote ApoB chylomicrons loading into COPII vesicles. Evidence supporting this is that the lack of MTP leads to ApoB accumulation at the ER. It is not known if secretion of other Tango1 cargos like collagen or Dpy also depends on MTP and PDI, but PDI is known also to form a complex with the collagen-modifying enzyme prolyl 4-hydroxylase. Previous work has shown that terminal cells lacking MTP show air-filling defects and fail to secrete Pio and Uninflatable to the apical membrane, and that loss of MTP in fat body cells also affects lipoprotein secretion, as it does in vertebrates. Since cells lacking MTP or Tango1 have similar phenotypes, it is plausible that the MTP function might be connected to the activity of Tango1 (Rios-Barrera, 2017).
The data is interpreted to mean that in the absence of Tango1, primary cargo accumulates in the ER, and in addition, there are secondary, indirect effects that can be suppressed by reducing the Tango1 cargo that overloads the ER. The secondary effects include activation of the ER stress response and intracellular accumulation of other trafficked proteins like Crb, laminins, and overexpressed proteins and probably also the accumulation of heterologous proteins like secreted HRP or GFP in other systems (Rios-Barrera, 2017).
The data suggest that primary and secondary cargo reach the ERES but fail to be trafficked further along the secretory pathway. In this model, primary cargo, probably recruited by adaptors, would be competing with other secondary cargo for ERES/COPII availability, creating a bottleneck at the ERES. This is consistent with recent experiments that show that in tango1-knockdown HeLa cells, VSVG-GFP trafficking does not stop completely, but is delayed. Furthermore, in these experiments, VSVG-GFP is mostly seen in association with Sec16 and Sec31, supporting the clogging model (Rios-Barrera, 2017).
It is not immediately clear why cargo accumulation in terminal cells lacking Tango1 affects the secretion of Crb but not of βPS integrin. While steady states are looked at in this analyses, Maeda (2016) measured the dynamics of secretion and found that loss of Tango1 leads to a reduced rate of secretion of VSVG-GFP, an effect that would have been missed for any proteins the current study classified as not affected by loss of Tango1. Irrespective, a range of mechanisms can be thought of that might be responsible for this difference, including alternative secretion pathways and differences in protein recycling. Alternative independent secretory pathways have been reported in different contexts. For instance, while both αPS1 and βPS integrin chains depend on Sec16 for their transport, the αPS1 chain can bypass the Golgi apparatus and can instead use the dGRASP-dependent pathway for its transport. It would be possible then that in terminal cells, βPS integrin is also trafficked through an alternative pathway that is not affected by loss of Tango1. Similarly, tracheal cells lacking Sec24-CD accumulate Gasp, Vermiform, and Fasciclin III, but not Crb, supporting a role for alternative secretion pathways for different proteins, as has been already proposed. Following this logic, overexpressed βPS integrin would then also be trafficked through a different route from that of the endogenous βPS integrin, possibly because of higher expression levels or because of the presence of the Venus fused to the normal protein (Rios-Barrera, 2017).
Drosophila Tango1 was initially found to facilitate collagen secretion in the fat body. More recently, the accumulation of other nonbulky proteins at the ER in the absence of Tango1 has led to the proposal of two models to explain these results: one in which Tango1 regulates general secretion, and the second one where Tango1 is specialized on the secretion of ECM components, since loss of Tango1 leads to the accumulation of the ECM molecules SPARC and collagen. The current results suggest a third explanation, where cargo accumulation in the ER might not necessarily be a direct consequence of only the loss of Tango1. Instead, in addition to depending on Tango1, some proteins of the ECM appear also to depend on each other for their efficient secretion. This is the case for laminins LanB1 and LanB2, which require trimerization before exiting the ER, while LanA can be secreted as a monomer. Loss of collagen itself leads to the intracellular accumulation of ECM components in fat body cells, such as the laminins and SPARC. Conversely, SPARC is required for proper collagen and laminin secretion and assembly in the ECM. Furthermore, intricate biochemical interactions take place between ECM components. Hence, due to the complex genetic and biochemical interactions between ECM components, the dependence of any one of them on Tango1 is difficult to determine without further biochemical evidence. The concept of interdependent protein transport from the ER as such is not new, as it has also been observed in other systems, for instance in immune complexes. During the assembly of T-cell receptor complexes and IgM antibodies, subunits that are not assembled are retained in the ER and degraded (Rios-Barrera, 2017).
Nevertheless, these observations in glial cells, which express laminins but not collagen, allow at least these requirements to be partly separate. This study found that laminins are accumulated due to general ER clogging and not because they rely on Tango1 for their export. This is based on observations that once the protein causing the ER block is removed, laminin secretion can continue in the absence of Tango1. It is still unclear why glial cells can secrete laminins in the absence of collagen whereas fat body cells cannot, but presumably laminin secretion can be mediated by different, unidentified cargo receptors expressed in glial cells (Rios-Barrera, 2017).
This study found that Sec16 forms aberrant aggregates in cells lacking Tango1, as in mammalian cell lines, and that the number of Sec16 particles is reduced. Other studies have shown that Tango1 overexpression produces larger ERES, and that Tango1 and Sec16 depend on each other for localization to ERES. In addition, lack of Tango1 also affects the distribution of Golgi markers. Thus, Tango1 influences not only the trafficking of cargos, but also the morphology of the secretory system (Rios-Barrera, 2017).
It had been suggested that the disorganization of ER and Golgi apparatus in cells lacking Tango1 might be an indirect consequence of the accumulation of Tango1 cargo. The work of Maeda (2016) has provided a possible explanation for the molecular basis, and proposed that Tango1 makes general secretion more efficient, but it has not formally excluded the possibility that the primary cause for the observed defects is secretory protein overload. This study has now shown that this is not the case: In the absence of Tango1, an aberrant ER and Golgi morphology is still observed even after the main primary substrates of Tango1 were removed and, thereby, secretion of other molecules was restored and the ER stress response was prevented (Rios-Barrera, 2017).
ERGIC53 accumulates at the ERES in the absence of Tango1, and this can be partly reversed by removing dpy. This is in apparent contradiction to findings in mammalian cells where Tango1 was necessary for the recruitment of membranes containing ERGIC53 to the budding collagen megacarrier vesicle. However, ERGIC53 also has Tango1-independent means of reaching the ER. The current results indicate that its retrieval from the ER to the ERGIC compartment depends directly or indirectly partly on Tango1. As a cargo receptor for glycoproteins, ERGIC53 may be retained at the ERES as a consequence of the accumulation of its own cargo at these sites. This would mean that it cannot be delivered back to the ERGIC or the cis-Golgi apparatus for further rounds of retrograde transport, which may, in turn, be an explanation for the enlargement of the Golgi matrix protein 130 kD (GM130) compartment seen after Tango1 knockdown (Rios-Barrera, 2017).
The finding that Tango1-depleted cells have a functional secretory pathway despite the ER-Golgi disorganization was unexpected. Stress stimuli like amino acid starvation (but not ER stress response itself) lead to Sec16 translocation into Sec bodies and inhibition of protein secretion. However, uncoupling of ER-Golgi organization from functional secretion has also been observed in other contexts. Loss of Sec23 or Sec24-CD leads to a target peptide sequence KDEL appearing in aggregates of varying sizes and intensities similar to those observed for Sec16 and for KDEL-RFP in cells lacking tango1. Also, GM130 is reduced in Sec23 mutant embryos. Despite these structural problems, these embryos do not show generalized secretion defects and also do not affect the functionality of the Golgi apparatus, as determined by glycosylation status of membrane proteins (Rios-Barrera, 2017).
Thus, Tango1 appears to have an important structural function in coordinating the organization of the ER and the Golgi apparatus, and this, in turn, may enhance vesicle trafficking. This fits with the role of Tango1 in recruiting ERGIC membranes to the ERES, and also with the effects of loss of Tango1 in the distribution of ER and Golgi markers. It has been proposed that the ER and Golgi apparatus in insects, which unlike in mammalian cells is not centralized but spread throughout the cytoplasm, is less efficient for secretion of bulky cargo than mammalian cells that can accommodate and transport it more efficiently through the Golgi ribbon. This difference could explain why tango1 knockout mice seem to have only collagen secretion defects and die only as neonates. However, a complete blockage of the ER might also be prevented by the activity of other MIA3/cTAGE5 family homologs. In mammalian cell culture experiments, even if loss of tango1 affects secretion of HRP, the secretion of other overexpressed molecules like alkaline phosphatase is not affected. This could also be because of the presence of other MIA3/cTAGE5 family homologs. By contrast, because there are no other MIA3/cTAGE5 family proteins in Drosophila, loss of tango1 may lead to the accumulation of a wider range of overexpressed proteins and more overt mutant phenotypes than in mammals (Rios-Barrera, 2017).
How tissues acquire their characteristic shape is a fundamental unresolved question in biology. While genes have been characterized that control local mechanical forces to elongate epithelial tissues, genes controlling global forces in epithelia have yet to be identified. This study describes a genetic pathway that shapes appendages in Drosophila by defining the pattern of global tensile forces in the tissue. In the appendages, shape arises from tension generated by cell constriction and localized anchorage of the epithelium to the cuticle via the apical extracellular-matrix protein Dumpy (Dp). Altering Dp expression in the developing wing results in predictable changes in wing shape that can be simulated by a computational model that incorporates only tissue contraction and localized anchorage. Three other wing shape genes, narrow, tapered, and lanceolate, encode components of a pathway that modulates Dp distribution in the wing to refine the global force pattern and thus wing shape (Ray, 2015).
This study has identified a group of genes that define the global force patterns that shape the appendages in Drosophila. During pupal development, shape is determined by a general contraction of the tissue in combination with localized anchorage to the pupal cuticle, which is mediated by the apical extracellular matrix (aECM) protein Dp. In the developing wing, Dp is localized to the wing margin such that, as tissue contraction proceeds, tension along the P-D axis elongates the wing and also draws the two wing surfaces together. Indeed, manipulating the pattern of Dp localization at this stage leads to dramatic changes in wing shape that reflect the underlying change in tissue anchorage. In the legs and antennae, Dp is found in a dense plaque at the distal tip of the appendage, and, as in the wing, tissue contraction results in tapering and elongation of the structure. Thus, this study has identified a genetic mechanism that determines shape by regulating the pattern of global tensile forces that the epithelium experiences during tissue contraction (Ray, 2015).
While the mechanism that this study has uncovered is clearly important for proper anchorage of the wing epithelium to the cuticle, it is only one part of the regulatory mechanism that leads to the localized attachment. Indeed, in a nw mutant, while the distribution of Dp is altered, it is still localized to the margin, thus other inputs must be involved in defining where Dp is localized during pupal development. In the wing, the localization of Dp to the margin is reminiscent of the expression of genes controlled by the Notch and Wingless pathways that define the dorsal-ventral compartment boundary. Indeed, the gene Dll is a downstream target of Wg, and knocking down dp in the cells that express Dll phenocopies the dp loss-of-function phenotype. Moreover, the notching associated with mutations in the Notch and Wg pathways, as well as their targets such as cut, are, in essence, defects in the anchorage of the margin during pupal development: the failure to specify the margin results in a gap in the expression of Dp which produces a phenotype not unlike that which is observed in dpp-Gal4>dpRNAi. Consistent with this, it has previously been shown that the notching associated with cut arises during pupal development during the period of hinge contraction. Thus, it may be that these phenotypes arise from a failure to localize Dp to specific regions of the margin rather than to cell death, as has been suggested previously (Ray, 2015).
In the leg and antenna, the localization of Dp to the extreme distal tip of the appendage is presumably under control of the P-D patterning system that operates in these tissues. Indeed, as in the wing, this study hasa shown that the retraction of the leg and antenna is produced by knocking down dp with Dll-Gal4, which is expressed in the most distal segments of both appendages. Moreover, the observed phenotypes are also associated with mutations in genes that affect specification of the distal most tarsal segments. Classical loss-of-function mutations of the Paired-type homeodomain protein Aristaless result in the same shortening of the arista as is observed with dp-RNAi. Similarly, loss-of-function alleles of the transcription factor Lim1 result in a shortening of the arista and the distal leg segments, which presumably reflects the failure to anchor the distal tip of the appendage to the pupal cuticle. Thus, it is speculated that for wings, legs, and antennae, the localized function of Dp appears to depend on cues from the developmental programs that pattern the appendages (Ray, 2015).
While Dp localization clearly depends on the positional cues set down in the imaginal discs, how these signaling molecules and transcription factors result in a localized pattern of Dp protein in the pupa remains unclear. Previous studies have shown that the dp mRNA is expressed throughout the developing wing in early pupal development (Wilkin, 2000), suggesting that the localized pattern of Dp expression that is evident at 18 hr APF results by a post-transcriptional mechanism. Throughout development, Dp functions as a link between the ectodermal epithelium and the cuticle, and, in order for the animal to molt, this connection must be periodically broken. During larval development, molting (or ecdysis) is initiated by apolysis, a process that separates epidermal cells from the old cuticle by secretion of a complex mixture of chitinases and proteases that degrade the carbohydrate and protein components of the exoskeleton, respectively. Given that Dp is essential for the link between epidermis and cuticle, it is a key target of the apolytic machinery. In the pupa, apolysis of the pupal cuticle occurs just prior to the onset of tissue contraction and is a prerequisite for generating global forces by differential anchorage. Given this, the localization of Dp and the localized anchorage of the appendage tissues to the cuticle presumably arise by protection of Dp at the margin and its degradation throughout the rest of the wing blade. These results suggest that in the wing, Nw acts to extend this protection more proximally to give rise to the wild-type shape of the wing (Ray, 2015).
The results show that coordination of the behavior of thousands of individual cells in the pupal wing is achieved by the precise regulation of global extrinsic forces that determine appendage shape. In this system, the wing hinge undergoes apical constriction to generate a pulling force that is transmitted through the tissue. As shown in this study, resistance to this force is provided by anchorage of the wing margin to the overlying pupal cuticle, resulting in anisotropic tension oriented predominantly along the P-D axis. As has been shown previously, cells respond to this tension via cell shape changes, oriented cell divisions, and cell rearrangements to drive tissue elongation and determine the final shape the wing. Thus, as a mechanical process, tissue shaping during pupal development depends on the magnitude of the force generated by the constricting cells, the strength of the anchorage resisting this force, and the fluidity of the tissue to relieve the tension (Ray, 2015).
Several lines of evidence suggest that these different components of the system operate toward an equilibrium state where opposing forces come to balance. For instance, if the force produced by the cell constriction exceeds the resistance, the anchorage ruptures, releasing the tissue. When this occurs, as in wings mutant for the hypomorphic allele dpov1, the margin collapses and the cells in the middle of the wing blade constrict further than they would otherwise. Similarly, the range of phenotypes observed in the interactions between nwD and dpov1 suggest that if the level of Dp falls below a certain threshold, the anchorage ruptures after hinge contraction has started, resulting in intermediate phenotypes that are flattened and retain some taper, but have nevertheless retracted. These observations suggest that cells in the wing blade actively respond to the pattern of tensile force with significant consequences on wing shape (Ray, 2015).
The data also indicate that hinge cells respond to the tension in the blade and constrict accordingly. Morphometric analysis of the nw mutant shows that the narrowing and extension of the wing blade is accompanied by a isometric contraction of the hinge, such that in the adult wing, the hinge is significantly smaller than in wild-type. Similarly, in the various dp mutants this study had examined, any change in the wing shape is accompanied by a corresponding change in hinge size and shape. The implication from these observations is that in the absence of sufficient resistance, hinge cells continue to undergo constriction, resulting in a corresponding change in hinge shape. Indeed, in the computational model, whenever the anchorage is released by any significant amount, the hinge is also found to contract further than in the wild-type simulation (Ray, 2015).
Taken together, these observations suggest that the system operates toward an equilibrium point where the force generated by hinge contraction is balanced with the resistance coming from the distal anchorage and the deformation properties of the cell matrix. Initially, the force generated by hinge contraction is offset by cell division and cell rearrangement, but when the cell division ceases, resistance in the epithelium feeds back on the hinge and eventually blocks further constriction. At this point, the system comes to equilibrium and the tension along the cell junctions equalizes. Notably, this model is consistent with previous reports, which have shown that after the initial phase of hinge contraction is completed, equalization of junction tension initiates the shift toward the hexagonal packing geometry that is characteristic of late stage pupal wings. The attractiveness of this kind of model is that it ensures robustness of the shaping mechanism and avoids the complications of tears or buckles that might arise from stochastic perturbations occurring during development. The nature of these feedbacks and how the equilibrium is achieved remain a question for further study (Ray, 2015).
As orthologs of Dp and Nw have been found, and in all sequenced insect genomes and in the crustacean Daphnia, yhr results suggest that the Nw-Dp system may be a key target for evolution of appendage shape in Arthropods. Indeed, given the tremendous variety of wing shapes that are found in insects, it is tempting to speculate that some of this variation is achieved by modulation of the Nw-Dp system, resulting in changes in the force patterns in the developing wing. For instance, in a recent report, wing shape differences in males of Nasonia vitripennis and Nasonia giraulti were attributed to differences in transcriptional regulation of an unpaired-like gene that was proposed to regulate proliferation in the developing wing. Significantly, the expression of this gene in both species was confined to the margin at the distal tip of the wing and strikingly prefigures the radius of curvature of the adult wing. Given these results, it is plausible that the difference in wing shape in these two species may arise from differences in anchorage of the wing tip, with the difference in cell number arising as a secondary consequence as in the case of nw. Similarly, the evolution of the diverse, specialized wing shapes in butterflies and moths has been attributed to defined expression of margin specific genes that prefigure the adult wing shape. While little is known of the elaboration of this prepattern during pupal development, the scalloping of the adult wing margin observed in many species—reminiscent of notching in the Drosophila wing—may well arise from precise deployment of the Nw-Dp system. Exploration of how Dp is regulated in different species may shed light on how the variety of insect wing shapes has evolved (Ray, 2015).
In all metazoans, assembly of the aECM is dependent on proteins that contain a common protein motif, the Zona Pellucida (ZP) domain, which is thought to act as a polymerization module promoting the formation of homo and heterotypic filaments. Genetic studies have implicated ZP-domain proteins in a variety of morphogenetic processes that typically involve shaping or remodeling of the apical domain of the cells. In Drosophila, ZP-domain proteins shape the embryonic denticles and hairs and the wing trichomes that form on the apical surface of the cells, and in C. elegans, the Cuticlulins are ZP-domain proteins involved in the formation of the alae. Similarly, in flies and nematodes, aECM proteins have been implicated as anchors in the cellular morphogenesis of sensory neurons, with Drosophila NompA functioning to anchor neural dendrites to the cuticular structures in sensory organs while in the nematode, the aECM proteins DEX-1 and DYF-7 anchor the dendritic tips during cell body migration to shape the amphid sense organs. Among these examples, these findings represent a different paradigm for how aECM proteins can influence morphogenesis: rather than affecting the behavior of individual cells, and consequently the shape of the tissue, the Nw-Dp mechanism defines global force patterns across the tissue to which the individual cells respond to give rise to appendage shape (Ray, 2015).
In vertebrates, the most well studied ZP-domain proteins are the eponymous zona pellucida proteins that form the extracellular coat of mammalian ova and the α- and β-tectorins that are required for the formation of the tectorial membrane in the ear. However, despite their importance for fertility and hearing, respectively, these proteins do not affect morphogenesis of the tissue per se. On the other hand, in the human kidney, the ZP-domain protein hensin/DMPT1 regulates morphogenesis of α- and β-Intercalated cells in the collecting tubules, and results from at least one study have found that global deletion of hensin results in embryonic lethality, suggesting a more general role in epithelial differentiation. Thus, there is mounting evidence from both invertebrates and vertebrates that the aECM plays an important role in morphogenesis and further studies on aECM proteins will undoubtedly reveal other roles they play in development and disease (Ray, 2015).
The Drosophila dumpy gene consists of seventy eight coding exons and encodes a huge extracellular matrix protein containing large numbers of epidermal growth factor-like (EGF) modules and a novel module called dumpy (DPY). A molecular analysis of forty five mutations in the dumpy gene of Drosophila melanogaster was carried out. Mutations in this gene affect three phenotypes: wing shape, thoracic cuticular defects, and lethality. Most of the mutations were chemically induced in a single dumpy allele and were analyzed using a nuclease that cleaves single base pair mismatches in reannealed duplexes followed by dHPLC. Additionally, several spontaneous mutations were analyzed. Virtually all of the chemically induced mutations, except for several in a single exon, either generate nonsense codons or lesions that result in downstream stop codons in the reading frame. The remaining chemically induced mutations remove splice sites in the nascent dumpy message. It is proposed that the vast majority of nonsense mutations that affect all three basic dumpy phenotypes are in constitutive exons, whereas nonsense mutants that remove only one or two of the basic functions are in alternatively spliced exons. Evolutionary comparisons of the dumpy gene from seven Drosophila species show strong conservation of the 5' ends of exons where mutants with partial dumpy function are found. In addition, reverse transcription PCR analyses reveal transcripts in which exons marked by nonsense mutations with partial dumpy function are absent (Carmon, 2010).
The complex Drosophila dumpy gene encodes a gigantic protein located in the apical extracellular matrix of epithelial cells. It has been shown to interact with several proteins notably during embryonic tracheal development. This study examined Dumpy's interactions in vivo with mutations in 20 genes previously recovered in a screen for recessive lethals that generate blisters when somatic clones are produced by mitotic crossing over during wing development. Primarily using double mutants, both dominant effects of the wing blister mutants and the effects of blister mutant clones on dumpy expression were looked for. Sixteen of the mutants either suppressed or enhanced dumpy mutant phenotypes indicating the large Dumpy protein is a very important component of the epithelial extracellular matrix in the wing. Dumpy also interacts strongly with held out wings, which is involved in RNA localization and possibly alternative splicing (Carmon, 2010).
EGF-domain O-GlcNAc transferase (Eogt) modifies EGF repeats in proteins that transit the secretory pathway, including Dumpy and Notch. This paper shows that the Notch ligands Delta and Serrate are also substrates of Eogt, that mutation of a putative UDP-GlcNAc binding DXD motif greatly reduces enzyme activity, and that Eogt and the cytoplasmic O-GlcNAc transferase Ogt have distinct substrates in Drosophila larvae. Loss of Eogt is larval lethal and disrupts Dumpy functions, but does not obviously perturb Notch signaling. To identify novel genetic interactions with eogt, this study investigated dominant modification of wing blister formation caused by knock-down of eogt. Unexpectedly, heterozygosity for several members of the canonical Notch signaling pathway suppressed wing blister formation. And importantly, extensive genetic interactions with mutants in pyrimidine metabolism were identified. Removal of pyrimidine synthesis alleles suppressed wing blister formation, while removal of uracil catabolism alleles was synthetic lethal with eogt knock-down. Therefore, Eogt may regulate protein functions by O-GlcNAc modification of their EGF repeats, and cellular metabolism by affecting pyrimidine synthesis and catabolism. It is proposed that eogt knock-down in the wing leads to metabolic and signaling perturbations that increase cytosolic uracil levels, thereby causing wing blister formation (Muller, 2013).
The O-linked-N-acetylglucosamine (O-GlcNAc) modification of cytoplasmic and nuclear proteins regulates basic cellular functions and is involved in the aetiology of diabetes and neurodegeneration. This intracellular O-GlcNAcylation is catalyzed by a single O-GlcNAc transferase, OGT. This study reports a novel OGT, EOGT, responsible for extracellular O-GlcNAcylation. Although both OGT and EOGT are regulated by hexosamine flux, EOGT localizes to the lumen of the endoplasmic reticulum and transfers GlcNAc to epidermal growth factor-like domains in an OGT-independent manner. Loss of Eogt gives phenotypes similar to those caused by defects in the apical extracellular matrix. Dumpy (Dp), a membrane-anchored extracellular protein, is O-GlcNAcylated, and EOGT is required for Dp-dependent epithelial cell-matrix interactions. Thus, O-GlcNAcylation of secreted and membrane glycoproteins is a novel mediator of cell-cell or cell-matrix interactions at the cell surface (Sakaidani, 2015).
The PIGSFEAST (PF) exon of the Drosophila dumpy gene is undergoing concerted evolution by the process of unequal crossing over. This study has developed a long-range PCR-based assay to amplify the approximately 12 kb long exon which contains variable numbers of 303 or 306 nt long repeats in a tandem array. This procedure was adapted to mutation accumulation lines of Drosophila melanogaster established by M. Wayne and L. Higgins. Nine new repeat length variants were found in these lines allowing measurement of the rate of unequal crossing over in the PF exon. The rate, which for several reasons is an underestimate, is 7.05 x 10-4 exchanges per generation (Carmon, 2010).
This study has determined by reverse Southern analysis and direct sequence comparisons that most of the dumpy gene has evolved in the dipteran and other insect orders by purifying selection acting on amino acid replacements. One region, however, is evolving rapidly due to unequal crossing over and/or gene conversion. This region, called "PIGSFEAST," or PF, encodes in D. melanogaster 30-47 repeats of 102 amino acids rich in serines, threonines, and prolines. The processes of concerted evolution have been operating on all species of Drosophila examined to date, but that an adjacent region has expanded in Anopheles gambiae, Aedes aegypti, and Tribolium castaneum, while the PF repeats are reduced in size and number. In addition, processes of concerted evolution have radically altered the codon usage patterns in D. melanogaster, D. pseudoobscura, and D. virilis compared with the rest of the dumpy gene. The dumpy gene is expressed on the inner surface of the micropyle of the mature oocyte, and it is proposed that, as in the abalone system, concerted evolution may be involved in adaptive changes affecting Dumpy's possible role in sperm-egg recognition (Carmon, 2007).
The formation of branched epithelial networks is fundamental to the development of many organs, such as the lung, the kidney or the vasculature. Little is known about the mechanisms that control cell rearrangements during tubulogenesis and regulate the size of individual tubes. Recent studies indicate that whereas the basal surface of tube cells interacts with the surrounding tissues and helps to shape the ramification pattern of tubular organs, the apical surface has an important role in the regulation of tube diameter and tube growth. This study reports that two proteins, Piopio (Pio) and Dumpy (Dp), containing a zona pellucida (ZP) domain are essential for the generation of the interconnected tracheal network in Drosophila melanogaster. Pio is secreted apically, accumulates in the tracheal lumen and possibly interacts with Dp through the ZP domains. In the absence of Pio and Dp, multicellular tubes do not rearrange through cell elongation and cell intercalation to form narrow tubes with autocellular junctions; instead they are transformed into multicellular cysts, which leads to a severe disruption of the branched pattern. It is proposed that an extracellular matrix containing Pio and Dp provides a structural network in the luminal space, around which cell rearrangements can take place in an ordered fashion without losing interconnections. These results suggest that a similar structural role might be attributed to other ZP-domain proteins in the formation of different branched organs (Jazwinska, 2003).
Search PubMed for articles about Drosophila Dumpy
Adler, P. N., Sobala, L. F., Thom, D., Nagaraj, R. (2013). dusky-like is required to maintain the integrity and planar cell polarity of hairs during the development of the Drosophila wing. Dev Biol, 379(1):76-91 PubMed ID: 23623898
Carmon, A., Topbas, F., Baron, M., MacIntyre, R. J. (2010). dumpy interacts with a large number of genes in the developing wing of Drosophila melanogaster. Fly (Austin), 4(2):117-127 PubMed ID: 20473031
Carmon, A., Wilkin, M., Hassan, J., Baron, M., MacIntyre, R. (2007). Concerted evolution within the Drosophila dumpy gene. Genetics, 176(1):309-325 PubMed ID: 17237523
Carmon, A., Larson, M., Wayne, M., MacIntyre, R. (2010). The rate of unequal crossing over in the dumpy gene from Drosophila melanogaster. J Mol Evol, 70(3):260-265 PubMed ID: 20204610
Carmon, A., Guertin, M. J., Grushko, O., Marshall, B., MacIntyre, R. (2010). A molecular analysis of mutations at the complex dumpy locus in Drosophila melanogaster. PLoS One, 5(8):e12319 PubMed ID: 20811586
Chu, W. C. and Hayashi, S. (2021). Mechano-chemical enforcement of tendon apical ECM into nano-filaments during Drosophila flight muscle development. Curr Biol. PubMed ID: 33545042
Drees, L., Konigsmann, T., Jaspers, M. H. J., Pflanz, R., Riedel, D., Schuh, R. (2019). Conserved function of the matriptase-prostasin proteolytic cascade during epithelial morphogenesis. PLoS Genet, 15(1):e1007882 PubMed ID: 30601807
Drees, L., Schneider, S., Riedel, D., Schuh, R., Behr, M. (2023). The proteolysis of ZP proteins is essential to control cell membrane structure and integrity of developing tracheal tubes in Drosophila. Elife, 12 PubMed ID: 37872795
Fernandes, I., Chanut-Delalande, H., Ferrer, P., Latapie, Y., Waltzer, L., Affolter, M., Payre, F., Plaza, S. (2010). Zona pellucida domain proteins remodel the apical compartment for localized cell shape changes. Dev Cell, 18(1):64-76 PubMed ID: 20152178
Ghosh, N., Treisman, J. E. (2024). Apical cell expansion maintained by Dusky-like establishes a scaffold for corneal lens morphogenesis. Sci Adv, 10(34):eado4167 PubMed ID: 39167639
Glassford, W. J., Johnson, W. C., Dall, N. R., Smith, S. J., Liu, Y., Boll, W., Noll, M., Rebeiz, M. (2015). Co-option of an Ancestral Hox-Regulated Network Underlies a Recently Evolved Morphological Novelty. Dev Cell, 34(5):520-531 PubMed ID: 26343453
Lai, Y. J., Chang, H. H., Lai, H., Xu, Y., Shiao, F., Huang, N., Li, L., Lee, M. S., Johnson, M. D., Wang, J. K., Lin, C. Y. (2015). N-Glycan Branching Affects the Subcellular Distribution of and Inhibition of Matriptase by HAI-2/Placental Bikunin. PLoS One, 10(7):e0132163 PubMed ID: 26171609
Muller, R., Jenny, A., Stanley, P. (2013). The EGF repeat-specific O-GlcNAc-transferase Eogt interacts with notch signaling and pyrimidine metabolism pathways in Drosophila. PLoS One, 8(5):e62835 PubMed ID: 23671640
Nagaraj, R., Adler, P. N. (2012). Dusky-like functions as a Rab11 effector for the deposition of cuticle during Drosophila bristle development. Development, 139(5):906-916 PubMed ID: 22278919
Ray, R. P., Matamoro-Vidal, A., Ribeiro, P. S., Tapon, N., Houle, D., Salazar-Ciudad, I. and Thompson, B. J. (2015). Patterned anchorage to the apical extracellular matrix defines tissue shape in the developing appendages of Drosophila. Dev Cell [Epub ahead of print]. PubMed ID: 26190146
Rios-Barrera, L. D., Sigurbjornsdottir, S., Baer, M. and Leptin, M. (2017). Dual function for Tango1 in secretion of bulky cargo and in ER-Golgi morphology. Proc Natl Acad Sci U S A 114(48): E10389-E10398. PubMed ID: 29138315
Roch, F., Alonso, C. R., Akam, M. (2003). Drosophila miniature and dusky encode ZP proteins required for cytoskeletal reorganisation during wing morphogenesis. J Cell Sci, 116(Pt 7):1199-1207 PubMed ID: 12615963
Sakaidani, Y., Nomura, T., Matsuura, A., Ito, M., Suzuki, E., Murakami, K., Nadano, D., Matsuda, T., Furukawa, K., Okajima, T. (2011). O-linked-N-acetylglucosamine on extracellular protein domains mediates epithelial cell-matrix interactions. Nat Commun, 2:583 PubMed ID: 22158438
Smith, S. J., Davidson, L. A., Rebeiz, M. (2020). Evolutionary expansion of apical extracellular matrix is required for the elongation of cells in a novel structure. Elife, 9 PubMed ID: 32338602
Tsuboi, A., Fujimoto, K., Kondo, T. (2023). Spatiotemporal remodeling of extracellular matrix orients epithelial sheet folding. Sci Adv, 9(35):eadh2154 PubMed ID: 37656799
Ray, R. P., Matamoro-Vidal, A., Ribeiro, P. S., Tapon, N., Houle, D., Salazar-Ciudad, I. and Thompson, B. J. (2015). Patterned anchorage to the apical extracellular matrix defines tissue shape in the developing appendages of Drosophila. Dev Cell [Epub ahead of print]. PubMed ID: 26190146
Chen, K. F., Peschel, N., Zavodska, R., Sehadova, H., Stanewsky, R. (2011). QUASIMODO, a Novel GPI-anchored zona pellucida protein involved in light input to the Drosophila circadian clock. Curr Biol, 21(9):719-729 PubMed ID: 21530261
Jazwinska, A., Ribeiro, C. and Affolter, M. (2003). Epithelial tube morphogenesis during Drosophila tracheal development requires Piopio, a luminal ZP protein. Nat. Cell Biol. 5: 895-901. PubMed ID: 12973360
Kandasamy, S., Couto, K. and Thackeray, J. (2021). A docked mutation phenocopies dumpy oblique alleles via altered vesicle trafficking. PeerJ 9: e12175. PubMed ID: 34721959
Smith, S. J., Davidson, L. A., Rebeiz, M. (2020). Evolutionary expansion of apical extracellular matrix is required for the elongation of cells in a novel structure. Elife, 9 PubMed ID: 32338602
Stempfl, T., Vogel, M., Szabo, G., Wulbeck, C., Liu, J., Hall, J. C., Stanewsky, R. (2002). Identification of circadian-clock-regulated enhancers and genes of Drosophila melanogaster by transposon mobilization and luciferase reporting of cyclical gene expression. Genetics, 160(2):571-593 PubMed ID: 11861563
Tsuboi, A., Fujimoto, K., Kondo, T. (2023). Spatiotemporal remodeling of extracellular matrix orients epithelial sheet folding. Sci Adv, 9(35):eadh2154 PubMed ID: 37656799
Wilkin, M. B., Becker, M. N., Mulvey, D., Phan, I., Chao, A., Cooper, K., Chung, H. J., Campbell, I. D., Baron, M., MacIntyre, R. (2000). Drosophila dumpy is a gigantic extracellular protein required to maintain tension at epidermal-cuticle attachment sites. Curr Biol, 10(10):559-567 PubMed ID: 10837220
date revised: 2 September 2024
Home page: The
Interactive Fly © 2011 Thomas Brody, Ph.D.
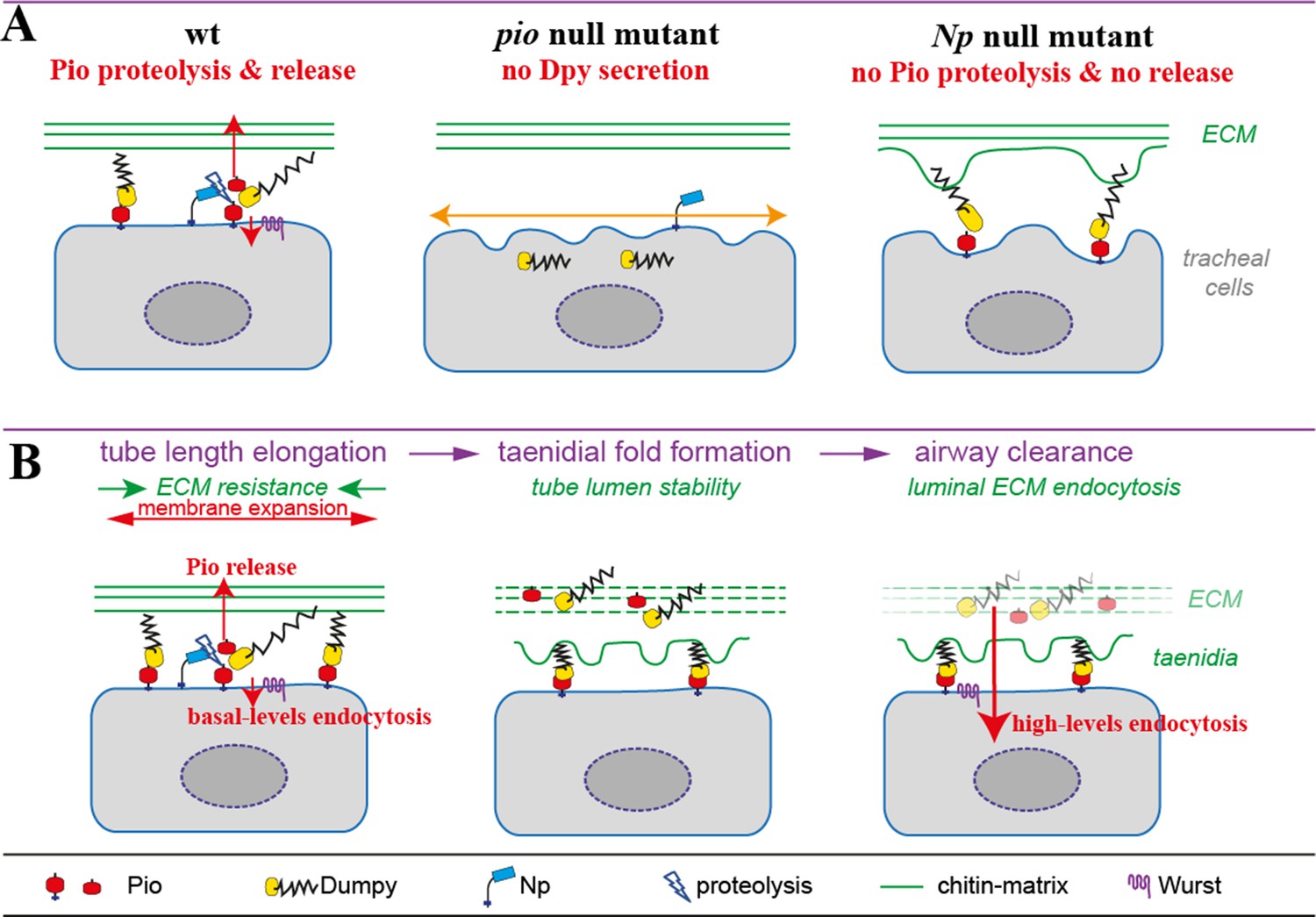{kind=link}