InteractiveFly: GeneBrief
piopio: Biological Overview | References
| Gene name - piopio Synonyms - Cytological map position - 60D7-60D8 Function - adhesion protein Keywords - mediates the adhesion of apical epithelial cell surfaces and the overlying apical extracellular matrix - interacts and cooperates with the ZP protein Dumpy at tracheal cells - undergoes ectodomain shedding by Notopleural - Dusky-like controls the arrangement of two zona pellucida domain proteins Dumpy and Piopio, external to the developing corneal lens. Loss of either dusky-like or dumpy delays chitin accumulation and disrupts the outer surface of the corneal lens |
Symbol - pio
FlyBase ID: FBgn0020521 Genetic map position - chr2R:24,587,707-24,597,862 NCBI classification - Zona pellucida (ZP) domain Cellular location - secreted |
| Recent literature | Drees, L., Schneider, S., Riedel, D., Schuh, R., Behr, M. (2023). The proteolysis of ZP proteins is essential to control cell membrane structure and integrity of developing tracheal tubes in Drosophila. Elife, 12 PubMed ID: 37872795
Summary: Membrane expansion integrates multiple forces to mediate precise tube growth and network formation. Defects lead to deformations, as found in diseases such as polycystic kidney diseases, aortic aneurysms, stenosis, and tortuosity.This study identified a mechanism of sensing and responding to the membrane-driven expansion of tracheal tubes. The apical membrane is anchored to the apical extracellular matrix (aECM) and causes expansion forces that elongate the tracheal tubes. The aECM provides a mechanical tension that balances the resulting expansion forces, with Dumpy being an elastic molecule that modulates the mechanical stress on the matrix during tracheal tube expansion. The zona pellucida (ZP) domain protein Piopio was shown to interact and cooperates with the ZP protein Dumpy at tracheal cells. To resist shear stresses which arise during tube expansion, Piopio undergoes ectodomain shedding by the Matriptase homolog Notopleural, which releases Piopio-Dumpy-mediated linkages between membranes and extracellular matrix. Failure of this process leads to deformations of the apical membrane, tears the apical matrix, and impairs tubular network function. Conserved ectodomain shedding was shown of the human TGFβ type III receptor by Notopleural and the human Matriptase, providing novel findings for in-depth analysis of diseases caused by cell and tube shape changes. |
Zhang, L., Ge, R., Yang, Y., Chen, K., Li, C. (2024). The zona pellucida protein piopio regulates the metamorphosis and reproduction in Tribolium castaneum. Archives of insect biochemistry and physiology, 116(1):e22122 PubMed ID: 38783685
Summary: The zona pellucida domain protein Piopio (Pio) was reported to mediate only the adhesion of the apical epithelial surface and the overlying apical extracellular matrix in Drosophila melanogaster, but the developmental roles of Pio were poorly understood in insects. To address this issue, this study comprehensively analyzed the function of Pio in Tribolium castaneum. Phylogenetic analysis indicated that pio exhibited one-to-one orthologous relationship among insects. T. castaneum pio had a 1236-bp ORF and contained eight exons. During development pio was abundantly expressed from larva to adult and lowly expressed at the late stage of embryo and adult, while it had more transcripts in the head, epidermis, and gut but fewer in the fat body of late-stage larvae. Knockdown of pio inhibited the pupation, eclosion, and reproduction of T. castaneum. The expression of vitellogenin 1 (Vg1), Vg2, and Vg receptor (VgR) largely decreased in pio-silenced female adults. Silencing pio increased the 20-hydroxyecdysone titer by upregulating phantom and spo expression but decreased the juvenile hormone (JH) titer through downregulating JHAMT3 and promoting JHE, JHEH-r4, and JHDK transcription. These results suggested that Pio might regulate the metamorphosis and reproduction via modulating the ecdysone and JH metabolism in T. castaneum. This study found the novel roles of pio in insect metamorphosis and reproduction, and provided the new insights for analyzing other zona pellucida proteins functions in insects. |
Gopfert, M., Yang, J., Rabadiya, D., Riedel, D., Moussian, B., Behr, M. (2025). Exoskeletal cuticle proteins enable Drosophila locomotion. Acta Biomater, PubMed ID: 40412508
Summary: Exo- and Endoskeleton function enables muscle-mediated locomotion in animals. In mammals, the defective protein matrix of bones found in systematic skeletal disorders such as osteoporosis causes fractures and severe skeletal deformations under high muscle tension. This study identified an analogous mechanism for integrating muscle-mediated tension into the apical extracellular matrix (aECM) of the invertebrate body wall exoskeleton. Obstructor chitin-binding proteins, the chitin deacetylases, Chitinases, and the matrix-protecting proteins Knickkopf and Retroactive are epidermally expressed during late embryogenesis. Their control of forming epidermal chitinous structures protects the exoskeletal aECM from collapsing when embryos start moving and hatch as larvae. In a larval locomotion assay the functions were tested of these cuticle related genes. Gene mutations and knockdowns caused changes in normal movement behavior and lower the speed of larvae. Moreover, the transmembrane Zona Pellucida domain protein Piopio provides the adhesion between the epidermal apical membrane and the overlaying chitinous aECM in a matriptase-dependent manner. A failure of Piopio and chitin-associated proteins leads to exoskeletal deformations and detachment from the epidermal membrane, destabilizing muscle forces and impairing larval mobility. The data identifies a protein network that transforms the chitinous aECM into a stable exoskeleton that directly resists muscle impact at epidermal tendon cells, thereby serving locomotion. Demonstrating the importance of these proteins in producing aECM as a three-dimensional cuticular scaffold for exoskeletal function opens up opportunities for the development of biomimetic applications of synthetic materials. |
Membrane expansion integrates multiple forces to mediate precise tube growth and network formation. Defects lead to deformations, as found in diseases such as polycystic kidney diseases, aortic aneurysms, stenosis, and tortuosity. This study identified a mechanism of sensing and responding to the membrane-driven expansion of tracheal tubes. The apical membrane is anchored to the apical extracellular matrix (aECM) and causes expansion forces that elongate the tracheal tubes. The aECM provides a mechanical tension that balances the resulting expansion forces, with Dumpy being an elastic molecule that modulates the mechanical stress on the matrix during tracheal tube expansion. The zona pellucida (ZP) domain protein Piopio was shown to interact and cooperates with the ZP protein Dumpy at tracheal cells. To resist shear stresses which arise during tube expansion, Piopio undergoes ectodomain shedding by the Matriptase homolog Notopleural, which releases Piopio-Dumpy-mediated linkages between membranes and extracellular matrix. Failure of this process leads to deformations of the apical membrane, tears the apical matrix, and impairs tubular network function. Conserved ectodomain shedding was shown of the human TGFβ type III receptor by Notopleural and the human Matriptase, providing novel findings for in-depth analysis of diseases caused by cell and tube shape changes (Drees, 2023).
Tracheal tube lumen expansion requires mechanical stress regulation at apical cell membranes and attached aECM. This involves the proteolytic processing of proteins that set local membrane-matrix linkages. Thus, the membrane microenvironment exhibits critical roles in regulating tube and network functionality (Drees, 2023).
ZP domain proteins organize protective aECM in the kidney, tectorial inner ear, and ZP, as well as in Drosophila epidermis, tendon cells, and appendages. Drosophila ZP domain proteins link the aECM to actin and polarity complexes in epithelial cells. Dumpy establishes force-resistant filaments for anchoring tendon cells to the pupal cuticle (Drees, 2023).
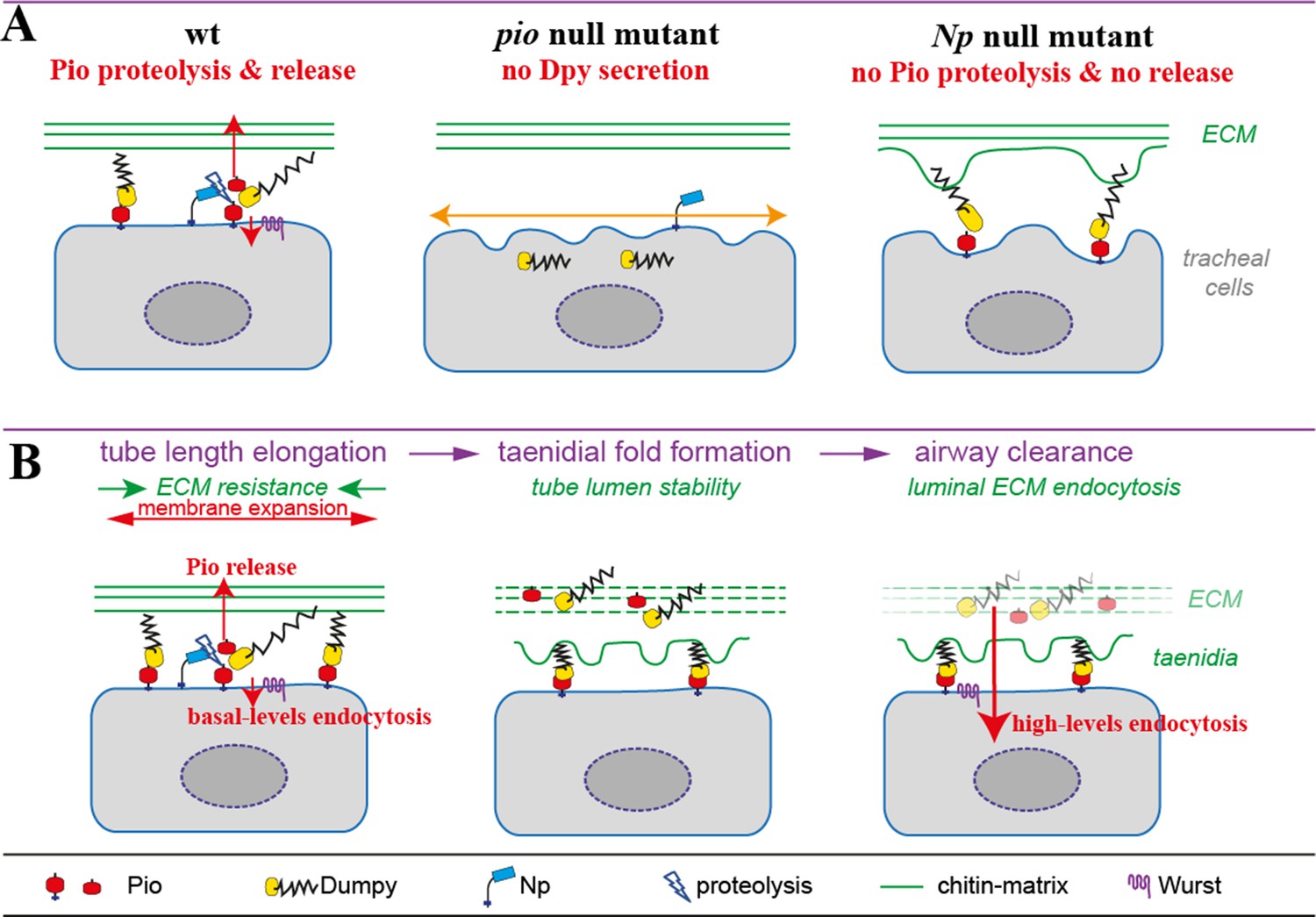
This study found that ZP protein-mediated microenvironmental changes increase the flexibility of membrane-matrix association, resulting from the activity of ZP domain proteins. Shear stress stimulates the activity of membrane-anchored proteases and potentially also apical transmembrane protease Np since the misdistribution of the tracheal cytoskeleton was not observed when blisters arise. The dynamic membrane-matrix association control is based on findings that loss of Np prevents Pio ectodomain shedding at the apical cell membrane resulting in immobile localization of Pio at the membrane and Dpy localization within the matrix. Direct interaction and overlapping subcellular localization at the cell surface showed that both proteins form a ZP matrix that potentially attaches membrane and ECM. Deregulation of Pio shedding blocks ZP matrix rearrangement and release of membrane-matrix linkages under tube expansion and subsequent shear stress. This destabilizes the microenvironment of membranes, causing blister formation at the membrane due to ongoing membrane expansion. Additionally, Pio could be part of a force-sensing signal transduction system destabilizing the membrane and matrix. The observation that the membrane deformations are maintained in Np mutant embryos supports the postulated Np function to redistribute and deregulate membrane-matrix associations in stage 16 embryos when tracheal tube length expands. In contrast, Np overexpression potentially uncouples the Pio-Dpy ZP matrix membrane linkages resulting very likely in unbalanced forces causing sinusoidal tubes (Drees, 2023).
The membrane defects observed in both Pio and Np mutants indicate errors in the coupling of the membrane matrix due to the involvement of Pio. In pio mutants, gaps appear between the deformed membrane and the apical matrix. These changes in apical cell membrane shape are consistent with increased cell and tube elongation in pio mutant embryos because the matrix is uncoupled from the membrane in such mutants. In contrast to pio mutants, the large membrane bulges in Np mutants affect the membrane and the apical matrix. Since apical Pio is not cleaved in Np mutants, the matrix is not uncoupled from the membrane as in pio mutant embryos but is likely more intensely coupled, which leads to tearing of the matrix axially along the membrane bulges, when the tube expands in length. If apical Pio detachment reduces coupling between the matrix and apical membrane, then it is likely that Np mutant embryos may exhibit a reduced tube length phenotype. In Np mutant embryos, average tracheal dorsal trunk length tends to be reduced compared to wt embryos, suggesting that Pio shedding is critical in controlling tracheal tube lumen length (Drees, 2023).
Is Pio ectodomain shedding in response to tension? Tension was not measured directly. However, the developmental profile of mechanical tension during tracheal tube length elongation in stage 16 embryos is consistent with the profile of Pio shedding. Np cleaves apical Pio during stage 16 when tube length expands. In contrast, Pio shedding decreases sharply at early stage 17 when tube elongation is completed. The model, therefore, predicts that loss of Pio or increased Pio secretion at stage 16 may reduce the coupling of the membrane matrix so that increased tracheal tube elongation is maintained until the end of stage 16, which is found in pio mutants and upon Np overexpression. Unknown proteases may likely be involved in Pio processing since cleaved mCherry::Pio is also detectable in inactive NpS990A cells. Previously a mutation at the Pio ZP domain (R196A) resistant to NP cleavage in cell culture experiments (Drees, 2019). Establishing a corresponding mutant fly line would be essential in determining whether the observed phenotype resembles the phenotype of the Np mutant embryos. In addition, unknown mechanisms, such as distinct membrane connections during development and emerging links to the developing cuticle, may also influence tension at the apical membrane during tube length control (Drees, 2023).
Indeed, the anti-Pio antibody, which detects all different Pio variants, showed a punctuate Pio pattern overlapping with the apical cell membrane markers Crb and Uninflatable (Uif) at the dorsal trunk cells of stage 16 embryos. Additionally, Pio antibody also revealed early tracheal expression from embryonic stage 11 onward, and due to Pio function in narrow dorsal and ventral branches, strong luminal Pio antibody staining is detectable from early stage 14 until stage 17, when airway protein clearance removes luminal contents. In the pio5m and pio17c mutants, Pio stainings were strongly reduced although some puncta were still detectable in the trachea. Similarly, Pio antibody staining is intracellular in the trachea of stage 11 pio2R-16 point mutation embryos. Interestingly, also dpy mutants showed strongly reduced and intracellular Pio antibody staining (Drees, 2023).
mCherry::Pio was generated as a tool for in vivo Pio expression and localization pattern analysis during tube lumen length expansion. The mCherry::Pio resembled the Pio antibody expression pattern from early tracheal development onward. However, luminal mCherry::Pio enrichment occurs specifically during stage 16, when tubes expand. The stage 16 embryos showed mCherry::Pio puncta accumulating apically in dorsal trunk cells. Moreover, mCherry::Pio puncta partially overlapped with Dpy::YFP and chitin at the taenidial folds, forming at apical cell membranes. Supported by several observations, such as antibody staining, video monitoring, FRAP experiments, and western blot studies, these findings indicate that Pio may play a significant role at the apical cell membrane and matrix in dorsal trunk cells of stage 16 embryo (Drees, 2023).
Furthermore, it was shown that Np mediates Pio ZP domain cleavage for luminal release of the short Pio variant during ongoing tube length expansion. The luminal cleaved mCherry::Pio is enriched at the end of stage 16 and finally internalized by the subsequent airway clearance process during stage 17 after tube length expansion. Such rapid luminal Pio internalization is consistent with a sharp pulse of endocytosis rapidly internalizing the luminal contents during stage 17. Wurst is required to mediate the internalization of proteins in the airways. Consistencly, during stage 17, luminal Pio antibody staining fades in control embryos but not in Wurst deficient embryos (Drees, 2023).
Nevertheless, Pio and its endocytosis depend on its interaction with the chitin matrix and the Np-mediated cleavage. In stage 16 wurst and mega mutant embryos, Pio antibody staining at the chitin cable, suggesting that Pio is cleaved and released into the dorsal trunk tube lumen. Also, the Cht2 overexpression did not prevent the luminal release of Pio. However, reduced wurst, mega function, and Cht2 overexpression caused an enrichment of punctuate Pio staining at the apical cell membrane and matrix . Although the three proteins are involved in different subcellular requirements, they all contribute to the determination of tube size by affecting either the apical cell membrane or the formation of a well-structured apical extracellular chitin matrix, indicating that changes at the apical cell membrane and matrix in stage 16 embryos affect the Pio pattern at the membrane. It also shows that local Pio linkages at the cell membrane and matrix are still cleaved by the Np function for luminal Pio release, which explains why those mutant embryos do not show pio mutant-like membrane deformations and Np-mutant-like bulges. This is in line with observations that tracheal Pio overexpression cannot cause tube size defects as the Np function is sufficient to organize local Pio linkages at the membrane and matrix. Therefore, it is unlikely that tracheal tube length defects in wurst and mega mutants as well as in Cht2 misexpression embryos are caused by the apical Pio density enrichment (Drees, 2023).
Nevertheless, oversized tube length due to the misregulation of the apical cell membrane and adjacent chitin matrix may cause changes to local Pio set linkages and the need for Np-mediated cleavage. Strikingly, a lack of Pio release was observed in Np mutants. This shows that Pio density at the membrane versus lumen depends predominantly on Np function. The molecular mechanisms that coordinate the Np-mediated Pio cleavage are unknown and will be necessary for understanding how tubes resist forces that impact cell membranes and matrices. On the other hand, Pio is required for the extracellular secretion of its interaction partner Dpy. At the same time, Dpy is needed for Pio localization at the cell membrane and its distribution into the tube lumen. Consistently, in vivo, mCherry::Pio, and Dpy::eYFP localization patterns overlap at the apical cell surface and within the tube lumen. These observations support the model that Pio and Dpy interact at the cell surface where Np mediates Pio cleavage to support luminal Pio release by the large and stretchable matrix protein Dpy (Drees, 2023).
Taenidial organization prevents the collapse of the tracheal tube. Therefore, cortical (apical) actin organizes into parallel-running bundles that proceed to the onset of cuticle secretion and correspond precisely to the cuticle’s taenidial folds. Mutant larvae of the F-actin nucleator formin DAAM show mosaic taenidial fold patterns, indicating a failure of alignment with each other and along the tracheal tubes. In contrast, pio mutant dorsal tracheal trunks contained increased ring spacing. Fusion cells are narrow doughnut-shaped cells where actin accumulates into a spotted pattern. Formins, such as Diaphanous, are essential in organizing the actin cytoskeleton. However, dorsal trunk tube fusion defects were not found as found in the presence of the activated Diaphanous (Drees, 2023).
On the other hand, ectopic expression of DAAM in fusion cells induces changes in apical actin organization but does not cause any phenotypic effects. DAAM is associated with the tyrosine kinase Src42A, which orients membrane growth in the axial tube dimension. Src42 overexpression elongates tracheal tubes due to flattened axially elongated dorsal trunk cells and AJ remodeling. Although flattened cells and tube overexpansion are similar in pio mutant embryos, mislocalization of AJ components was not observed, as found upon constitutive Src42 activation. Instead, an unusual stretched appearance of AJs was detected at the fusion cells of pio mutant dorsal trunks, which has not been observed before and may play a role in regulating axial taenidial fold spacing and tube elongation (Drees, 2023).
Self-organizing physical principles govern the regular spacing pattern of the tracheal taenidial folds. The actomyosin cortex and increased actin activity before and turnover at stage 16 drive the regular pattern formation. However, the cell cortex and actomyosin are in frictional contact with a rigid apical ECM. The Src42A mutant embryos contain shortened tube length but increased taenidial fold period pattern due to decreased friction. In contrast, the chitinase synthase mutant kkv1 has tube dilation defects and no regular but an aberrant pearling pattern caused by zero fiction (Drees, 2023).
In contrast, pio mutant embryos do not contain tube dilation defects or shortened tubes but increased tube length. Furthermore, chitin-binding probe (cbp) and antibody stainings reveal the presence of a luminal chitin cable and a solid aECM structure in pio mutant stage 16 embryos. In addition, apical actin enrichment in tracheal cells of pio mutant embryos appeared wt-like. Nonetheless, pio mutant embryos show an increased taenidial fold period compared with wt, indicating a decreased friction. Thus, it is proposed that the lack of Pio reduces friction. Reasons might be subtle defects of actomyosin constriction or chitin matrix, which was not detected in the pio mutant tracheal cells. Further reasons for lower friction might also be the loss of Pio set local linkages between apical cortex and aECM in stage 16 embryos, which are modified by Np, as proposed in in a model (see Model of apical Pio and Dpy matrix at the apical cell surface and Pio proteolysis and release. (Drees, 2023).
Heterozygous and homozygous pio mutant embryos generally do not show tubal collapse. However, the loss of Pio and accompanying lack of Dpy secretion in stage 17 pio mutant embryos led to the loss of a Pio/Dpy matrix, impacting the late embryonic maturation and differentiation of a normal chitin matrix at the apical cell surface. TEM images reveal reduced dense chitin matrix material at taenidial folds and misarranged taenidial fold pattern, suggesting impaired taenidial function prevents tube lumen from collapsing after tube protein clearance. wurst knockdown and mutant embryos do not show general tube collapse, but luminal chitin fiber organization is disturbed in stage 17 embryos. Therefore, transheterozygous wurst;pio mutant embryos may combine both defects and suffer from maturation deficits of the chitin/ZP matrix at the apical cell surface and within the tube lumen, which finally causes a high number of embryos with incomplete gas-filling due to tube collapse. These maturation deficits are even more dramatic in the wurst;pio double mutants, which show no gas-filling (Drees, 2023).
These studies on human Matriptase provide evidence for a mechanistic conservation of ZP domain protein as a substrate for ectodomain shedding. The upregulation of Matriptase activity and increased TGFβ receptor density affect human and mouse model idiopathic pulmonary fibrosis cells on pulmonary fibrogenesis. Furthermore, the human Matriptase induces the release of proinflammatory cytokines in endothelial cells, which contribute to atherosclerosis and probably also to abdominal aortic aneurysms. The membrane bulges arising in the Drosophila model during tracheal tube elongation upon Np loss of function showed analogy to the appearance of artery aneurysms. Bulges with varying phenotypic expression in different organs can lead to aortic rupture due to fragile artery walls or degeneration of layers in responses to stimuli, such as shear stresses. Indeed, aneurysm development is forced by alterations in the ECM and is characterized by extensive ECM fragmentation caused by shedding of membrane-bound proteins (Drees, 2023).
This study identified a dynamic control of matrix proteolysis, very likely enabling fast and site-specific uncoupling of membrane-matrix linkages when tubes expand. Such a scenario has not yet been studied in angiogenesis. It may represent a new starting point for genetic studies to decipher the putative roles of ZP domain proteins and Matriptase in clinically relevant syndromes, including the formation of aneurysms caused by membrane deformation and defects in size determination of airways and vessels (Drees, 2023).
Biological systems are inherently noisy; however, they produce highly stereotyped tissue morphology. Drosophila pupal wings show a highly stereotypic folding through uniform expansion and subsequent buckling of wing epithelium within a surrounding cuticle sac (see Emergence of stereotypic folding within a cuticle sac). The folding pattern produced by buckling is generally stochastic; it is thus unclear how buckling leads to stereotypic tissue folding of the wings. This study found that the extracellular matrix (ECM) protein, Dumpy, guides the position and direction of buckling-induced folds. Dumpy anchors the wing epithelium to the overlying cuticle at specific tissue positions. Tissue-wide alterations of Dumpy deposition and degradation yielded different buckling patterns. In summary, it is proposed that spatiotemporal ECM remodeling shapes stereotyped tissue folding through dynamic interactions between the epithelium and its external structures (Tsuboi, 2023).
This study has revealed that the spatiotemporally coordinated deposition and destruction of the ECM protein (Dpy) guide the position and direction of buckling. The results demonstrate that although cell populations show spatially homogeneous cellular behaviors (i.e., cell flattening), they can yield stereotypic tissue buckling morphology through the positional information encoded by ECM remodeling (Tsuboi, 2023).
Dpy, which anchors the wing tissue and cuticle, is constructed at specific positions (see Dumpy regulates the stereotypic buckling direction by connecting the wing epithelium and the pupal cuticle along veins). The position of Dpy anchorage is likely controlled posttranslationally, because Dpy overexpression did not result in the ectopic formation of Dpy anchorage. The construction of the Dpy matrix involves multiple processes, including synthesis, secretion, assembly into polymers, and association of the cuticle with the apical cell surface. Although the signaling pathway regulating Dpy anchoring remains unknown, the loss of vein Dpy in the mutant wing lacking veins suggests that positional information and signaling pathways related to wing vein patterning may contribute to the spatial differences in Dpy structure formation (Tsuboi, 2023).
Dpy degradation was found to initiate in the distal-posterior region, and the spatial pattern of degradation controls the position of the marginal fold (see Dumpy degradation is indispensable for the marginal fold formation). The degradation of Dpy is regulated by the action of Stubble (Sb) and Np apical transmembrane proteases, suggesting that the spatial pattern of Np/Sb expression or localization to the apical surface may control the degradation pattern of Dpy. Alternatively, another possible mechanism for the propagation pattern of Dpy degradation could be attributed to the spatial pattern of proteolytic activation of the proteases, because Np and Sb are expressed as catalytically inactive zymogens and require proteolytic cleavage to become active. In addition, regional differences in the physical properties of Dpy and its sensitivity to the proteases could contribute to the propagation pattern of Dpy degradation. It has been reported that Dpy undergoes filamentous conversion in response to increasing tension during indirect flight muscle development. Considering that Dpy filaments in wing tissue are formed under anisotropic proximal-distal–oriented tension caused by hinge contraction, the extent of Dpy filamentous conversion may vary regionally within the tissue depending on the applied stress. It would be interesting to investigate the regional difference in the physical properties of Dpy filaments within a tissue and their relevance to the propagation pattern of Dpy degradation (Tsuboi, 2023).
These findings are unique as they reveal the potential of external cues in generating stereotypic 3D tissue shapes within a spatially confined environment. This ECM-based mechanism should confer a potential means to generate diverse and controllable 3D tissue shaping in parallel with cell-intrinsic genetic programming. Future application of this ECM modification to tissue engineering would pave the way for manufacturing precisely folded tissues in any desired manner (Tsuboi, 2023).
Extracellular matrix (ECM) assembly and remodelling is critical during development and organ morphogenesis. Dysregulation of ECM is implicated in many pathogenic conditions, including cancer. The type II transmembrane serine protease matriptase and the serine protease prostasin are key factors in a proteolytic cascade that regulates epithelial ECM differentiation during development in vertebrates. This study shows by rescue experiments that the Drosophila proteases Notopleural (Np) and Tracheal-prostasin (Tpr) are functional homologues of matriptase and prostasin, respectively. Np mediates morphogenesis and remodelling of apical ECM during tracheal system development and is essential for maintenance of the transepithelial barrier function. Both Np and Tpr degrade the zona pellucida-domain (ZP-domain) protein Dumpy, a component of the transient tracheal apical ECM. Tpr zymogen and the ZP domain of the ECM protein Piopio are cleaved by Np and matriptase in vitro. These data indicate that the evolutionarily conserved ZP domain, present in many ECM proteins of vertebrates and invertebrates, is a novel target of the conserved matriptase-prostasin proteolytic cascade (Drees, 2019).
There is increasing evidence that tissue-specific modifications of basic cellular functions play an important role in development and disease. To identify the functions of COPI coatomer-mediated membrane trafficking in Drosophila development, this study aimed to create loss-of-function mutations in the gammaCOP gene, which encodes a subunit of the COPI coatomer complex. ammaCOP is essential for the viability of the Drosophila embryo. In the absence of zygotic gammaCOP activity, embryos die late in embryogenesis and display pronounced defects in morphogenesis of the embryonic epidermis and of tracheal tubes. The coordinated cell rearrangements and cell shape changes during tracheal tube morphogenesis critically depend on apical secretion of certain proteins. Investigation of tracheal morphogenesis in gammaCOP loss-of-function mutants revealed that several key proteins required for tracheal morphogenesis are not properly secreted into the apical lumen. As a consequence, gammaCOP mutants show defects in cell rearrangements during branch elongation, in tube dilation, as well as in tube fusion. Genetic evidence is presented that a specific subset of the tracheal defects in gammaCOP mutants is due to the reduced secretion of the Zona Pellucida protein Piopio. Thus, a critical target protein of COPI-dependent secretion in epithelial tube morphogenesis was identified. These studies highlight the role of COPI coatomer-mediated vesicle trafficking in both general and tissue-specific secretion in a multicellular organism. Although COPI coatomer is generally required for protein secretion, the phenotypic effect of gammaCOP mutations is surprisingly specific. Importantly, a distinct aspect of the gammaCOP phenotype is attributed to the effect on a specific key target protein (Grieder, 2008).
Adhesion between epithelial cells and extracellular substrates is normally mediated through basal adhesion complexes. However, some cells also possess comparable junctions on their apical surface. This study describes two new Drosophila proteins, Piopio and Papillote, that are required for the link between the apical epithelial surface and the overlying apical extracellular matrix (aECM). The two proteins share a zona pellucida (ZP) domain with mammalian aECM components, including the tectorins found in the vertebrate inner ear. Tagged versions of both proteins localized to the apical epithelial surface. Mutations in piopio, papillote and dumpy (another gene encoding a ZP-domain protein) cause defects in the innermost layer of the aECM and its detachment from the epidermis. Loss of Piopio, but not Papillote or Dumpy, causes the absence of specialized microtubule bundles from pupal wings, suggesting that Piopio plays a role in microtubule organization. Thus, ZP domain-containing proteins may have shared functions within the aECM, while also exhibiting specific interactions with the cytoskeleton (Bokel, 2005).
The formation of branched epithelial networks is fundamental to the development of many organs, such as the lung, the kidney or the vasculature. Little is known about the mechanisms that control cell rearrangements during tubulogenesis and regulate the size of individual tubes. Recent studies indicate that whereas the basal surface of tube cells interacts with the surrounding tissues and helps to shape the ramification pattern of tubular organs, the apical surface has an important role in the regulation of tube diameter and tube growth. This study reports that two proteins, Piopio (Pio) and Dumpy (Dp), containing a zona pellucida (ZP) domain are essential for the generation of the interconnected tracheal network in Drosophila melanogaster. Pio is secreted apically, accumulates in the tracheal lumen and possibly interacts with Dp through the ZP domains. In the absence of Pio and Dp, multicellular tubes do not rearrange through cell elongation and cell intercalation to form narrow tubes with autocellular junctions; instead they are transformed into multicellular cysts, which leads to a severe disruption of the branched pattern. It is proposed that an extracellular matrix containing Pio and Dp provides a structural network in the luminal space, around which cell rearrangements can take place in an ordered fashion without losing interconnections. These results suggest that a similar structural role might be attributed to other ZP-domain proteins in the formation of different branched organs (Jazwinska. 2003).
Integrins are evolutionarily conserved transmembrane alpha,beta heterodimeric receptors involved in cell-to-matrix and cell-to-cell adhesions. In Drosophila the position-specific (PS) integrins mediate the formation and maintenance of junctions between muscle and epidermis and between the two epidermal wing surfaces. Besides integrins, other proteins are implicated in integrin-dependent adhesion. In Drosophila, somatic clones of mutations in PS integrin genes disrupt adhesion between wing surfaces to produce wing blisters. To identify other genes whose products function in adhesion between wing surfaces, a screen was conducted for autosomal mutations that produce blisters in somatic wing clones. 76 independent mutations were isolated in 25 complementation groups, 15 of which contain more than one allele. Chromosomal sites were determined by deficiency mapping, and genetic interactions with mutations in the beta PS integrin gene myospheroid were investigated. Mutations in four known genes (blistered, Delta, dumpy and mastermind) were isolated. Mutations were isolated in three new genes (piopio, rhea and steamer duck) that affect myo-epidermal junctions or muscle function in embryos. Mutations in three other genes (kakapo, kiwi and moa) may also affect cell adhesion or muscle function at hatching. These new mutants provide valuable material for the study of integrin-dependent cell-to-cell adhesion (Prout, 1997).
Search PubMed for articles about Drosophila Piopio
Bokel, C., Prokop, A., Brown, N. H. (2005). Papillote and Piopio: Drosophila ZP-domain proteins required for cell adhesion to the apical extracellular matrix and microtubule organization. J Cell Sci, 118(Pt 3):633-642 PubMed ID: 15657084
Drees, L., Konigsmann, T., Jaspers, M. H. J., Pflanz, R., Riedel, D., Schuh, R. (2019). Conserved function of the matriptase-prostasin proteolytic cascade during epithelial morphogenesis. PLoS Genet, 15(1):e1007882 PubMed ID: 30601807
Drees, L., Schneider, S., Riedel, D., Schuh, R., Behr, M. (2023). The proteolysis of ZP proteins is essential to control cell membrane structure and integrity of developing tracheal tubes in Drosophila. Elife, 12 PubMed ID: 37872795
Grieder, N. C., Caussinus, E., Parker, D. S., Cadigan, K., Affolter, M., Luschnig, S. (2008). gammaCOP is required for apical protein secretion and epithelial morphogenesis in Drosophila melanogaster. PLoS One, 3(9):e3241 PubMed ID: 18802472
Jazwinska, A., Ribeiro, C. and Affolter, M. (2003). Epithelial tube morphogenesis during Drosophila tracheal development requires Piopio, a luminal ZP protein. Nat. Cell Biol. 5: 895-901. PubMed ID: 12973360
Tsuboi, A., Fujimoto, K., Kondo, T. (2023). Spatiotemporal remodeling of extracellular matrix orients epithelial sheet folding. Sci Adv, 9(35):eadh2154 PubMed ID: 37656799
date revised: 5 January 2026
Home page: The
Interactive Fly © 2026 Thomas Brody, Ph.D.
{kind=link}