InteractiveFly: GeneBrief
missing oocyte: Biological Overview | References
|
Gene name - missing oocyte
Synonyms - Cytological map position - 22D4-22D4 Function - conserved novel protein Keywords - regulates nuclear architecture and meiotic progression in early ovarian cysts, a novel interacting partner of the conserved nucleoporin Seh1, influences meiotic progression and oocyte fate |
Symbol - mio
FlyBase ID: FBgn0031399 Genetic map position - chr2L:2,217,363-2,220,625 Classification - WD40 repeat protein Cellular location - nuclear |
| Recent literature | Chng, W. A., Koch, R., Li, X., Kondo, S., Nagoshi, E. and Lemaitre, B. (2017). Transforming Growth Factor beta/Activin signaling in neurons increases susceptibility to starvation. PLoS One 12(10): e0187054. PubMed ID: 29084242
Summary: Animals rely on complex signaling network to mobilize its energy stores during starvation. Previous work has shown that the sugar-responsive TGFbeta/Activin pathway, activated through the TGFbeta ligand Dawdle, plays a central role in shaping the post-prandial digestive competence in the Drosophila midgut. Nevertheless, little is known about the TGFbeta/Activin signaling in sugar metabolism beyond the midgut. This study addresses the importance of Dawdle (Daw) after carbohydrate ingestion. Daw expression was found to be coupled to dietary glucose through the evolutionarily conserved Mio-Mlx transcriptional complex. In addition, Daw activates the TGFbeta/Activin signaling in neuronal populations to regulate triglyceride and glycogen catabolism and energy homeostasis. Loss of those neurons depleted metabolic reserves and rendered flies susceptible to starvation. |
Zhang, Y., Ting, C. Y., Yang, S., Reich, J., Fru, K. and Lilly, M. A. (2023). Wdr59 promotes or inhibits TORC1 activity depending on cellular context. Proc Natl Acad Sci U S A 120(1): e2212330120. PubMed ID: 36577058
Summary: Target of Rapamycin Complex I (TORC1) is a central regulator of metabolism in eukaryotes that responds to a wide array of negative and positive inputs. The GTPase-activating protein toward Rags (GATOR) signaling pathway acts upstream of TORC1 and is comprised of two subcomplexes. The trimeric GATOR1 complex inhibits TORC1 activity in response to amino acid limitation by serving as a GTPase-activating protein (GAP) for the TORC1 activator RagA/B, a component of the lysosomally located Rag GTPase. The multi-protein GATOR2 complex inhibits the activity of GATOR1 and thus promotes TORC1 activation. This study reports that Wdr59, originally assigned to the GATOR2 complex based on studies performed in tissue culture cells, unexpectedly has a dual function in TORC1 regulation in Drosophila. <../aimorph/oocyte.htm">ovary and the eye imaginal disc brain complex, Wdr59 inhibits TORC1 activity by opposing the GATOR2-dependent inhibition of GATOR1. Conversely, in the Drosophila fat body, Wdr59 promotes the accumulation of the GATOR2 component Mio and is required for TORC1 activation. Similarly, in mammalian HeLa cells, Wdr59 prevents the proteolytic destruction of GATOR2 proteins Mio and Wdr24. Consistent with the reduced levels of the TORC1-activating GATOR2 complex, Wdr59KOs HeLa cells have reduced TORC1 activity which is restored along with GATOR2 protein levels upon proteasome inhibition. Taken together, our data support the model that the Wdr59 component of the GATOR2 complex functions to promote or inhibit TORC1 activity depending on cellular context. |
Chng, W. A., Koch, R., Li, X., Kondo, S., Nagoshi, E. and Lemaitre, B. (2017). Transforming Growth Factor beta/Activin signaling in neurons increases susceptibility to starvation. PLoS One 12(10): e0187054. PubMed ID: 29084242
Summary: Animals rely on complex signaling network to mobilize its energy stores during starvation. Previous work has shown that the sugar-responsive TGFbeta/Activin pathway, activated through the TGFbeta ligand Dawdle, plays a central role in shaping the post-prandial digestive competence in the Drosophila midgut. Nevertheless, little is known about the TGFbeta/Activin signaling in sugar metabolism beyond the midgut. This study addresses the importance of Dawdle (Daw) after carbohydrate ingestion. Daw expression was found to be coupled to dietary glucose through the evolutionarily conserved Mio-Mlx transcriptional complex. In addition, Daw activates the TGFbeta/Activin signaling in neuronal populations to regulate triglyceride and glycogen catabolism and energy homeostasis. Loss of those neurons depleted metabolic reserves and rendered flies susceptible to starvation. |
Chng, W. A., Koch, R., Li, X., Kondo, S., Nagoshi, E. and Lemaitre, B. (2017). Transforming Growth Factor beta/Activin signaling in neurons increases susceptibility to starvation. PLoS One 12(10): e0187054. PubMed ID: 29084242
Summary: Animals rely on complex signaling network to mobilize its energy stores during starvation. Previous work has shown that the sugar-responsive TGFbeta/Activin pathway, activated through the TGFbeta ligand Dawdle, plays a central role in shaping the post-prandial digestive competence in the Drosophila midgut. Nevertheless, little is known about the TGFbeta/Activin signaling in sugar metabolism beyond the midgut. This study addresses the importance of Dawdle (Daw) after carbohydrate ingestion. Daw expression was found to be coupled to dietary glucose through the evolutionarily conserved Mio-Mlx transcriptional complex. In addition, Daw activates the TGFbeta/Activin signaling in neuronal populations to regulate triglyceride and glycogen catabolism and energy homeostasis. Loss of those neurons depleted metabolic reserves and rendered flies susceptible to starvation. |
In single-cell eukaryotes the pathways that monitor nutrient availability are central to initiating the meiotic program and gametogenesis. In Saccharomyces cerevisiae an essential step in the transition to the meiotic cycle is the down-regulation of the nutrient-sensitive target of rapamycin complex 1 (TORC1; see Drosophila Tor pathway) by the increased minichromosome loss 1/ GTPase-activating proteins toward Rags 1 (Iml1/GATOR1) complex in response to amino acid starvation. How metabolic inputs influence early meiotic progression and gametogenesis remains poorly understood in metazoans. This study defined opposing functions for the TORC1 regulatory complexes Iml1/GATOR1 and GATOR2 during Drosophila oogenesis. As is observed in yeast, the Iml1/GATOR1 complex inhibits TORC1 activity to slow cellular metabolism and drive the mitotic/meiotic transition in developing ovarian cysts. In iml1 germline depletions, ovarian cysts undergo an extra mitotic division before meiotic entry. The TORC1 inhibitor rapamycin can suppress this extra mitotic division. Thus, high TORC1 activity delays the mitotic/meiotic transition. Conversely, mutations in Tor, which encodes the catalytic subunit of the TORC1 complex, result in premature meiotic entry. Later in oogenesis, the GATOR2 components Missing oocyte (Mio)
In yeast, the inhibition of the nutrient-sensitive target of rapamycin complex 1 (TORC1) in response to amino acid limitation is essential for cells to transit from the mitotic cycle to the meiotic cycle. In response to amino acid starvation, the Iml1 complex, comprising the Iml1, Nitrogen permease regulator-like 2 (Npr2), and Nitrogen permease regulator-like 3 (Npr3) proteins in yeast and the respective orthologs DEPDC5, Nprl2, and Nprl3 in mammals, inhibits TORC1 activity. The Iml1 complex, which has been renamed the 'GTPase-activating proteins toward Rags 1' (GATOR1) complex in higher eukaryotes, functions as a GTPase-activating protein complex that inactivates RagsA/B or Gtr1 in mammals and yeast, respectively, thus preventing the activation of TORC1. In the yeast Saccharomyces cerevisiae, mutations in the Iml1 complex components Npr2 and Npr3 result in a failure to down-regulate TORC1 activity in response to amino acid starvation and block meiosis and sporulation. As is observed in yeast, in Drosophila, Nprl2 and Nprl3 mediate a critical response to amino acid starvation (Wei, 2014a). However, their roles in meiosis and gametogenesis remain unexplored (Wei, 2014b).
Recent reports indicate that the Iml1, Npr2, and Npr3 proteins are components of a large multiprotein complex originally named the 'Seh1-associated' (SEA) complex in budding yeast and the 'GATOR' complex in higher eukaryotes. The SEA/GATOR complex contains eight highly conserved proteins. The three proteins described above, Iml1/DEPDC5, Npr2/Nprl2, and Npr3/Nprl3, form the Iml1/GATOR1 complex and inhibit TORC1. The five remaining proteins in the complex, Seh1, Sec13, Sea4/Mio, Sea2/WDR24, and Sea3/WDR59, which have been designated the 'GATOR2' complex in multicellular organisms, oppose the activity of Iml1/GATOR1 and thus promote TORC1 activity (Wei, 2014b).
Little is known about the physiological and/or developmental requirements for the GATOR2 complex in multicellular organisms. However, in Drosophila the GATOR2 components Mio and Seh1 interact physically and genetically and exhibit strikingly similar ovarian phenotypes, with null mutations in both genes resulting in female sterility (Senger, 2011; Wei, 2014a). In Drosophila females, oocyte development takes place within the context of an interconnected germline syncytium, also referred to as an 'ovarian cyst'. Ovarian cyst formation begins at the tip of the germarium when a cystoblast, the daughter of a germline stem cell, undergoes four synchronous divisions with incomplete cytokinesis to produce 16 interconnected cells. Actin-stabilized cleavage furrows, called 'ring canals', connect cells within the cyst. Each 16-cell cyst develops with a single oocyte and 15 polyploid nurse cells which ultimately are encapsulated by a somatically derived layer of follicle cells to produce an egg chamber. Each ovary is comprised of ∼15 ovarioles that consist of a single germarium followed by a string of egg chambers in successively older stages of development. In mio- and seh1-mutant egg chambers, the oocyte enters the meiotic cycle, but as oogenesis proceeds, the oocyte fate and the meiotic cycle are not maintained stably (Senger, 2011; Wei, 2014a). Ultimately, a large fraction of mio and seh1 oocytes enter the endocycle and develop as polyploid nurse cells. A mechanistic understanding of how mio and seh1 influence meiotic progression and oocyte fate has remained elusive (Wei, 2014b).
This study demonstrates that the Iml1/GATOR1 complex down-regulates TORC1 activity to promote the mitotic/meiotic transition in Drosophila ovarian cysts. Depleting iml1 in the female germ line delays the mitotic/meiotic transition, so that ovarian cysts undergo an extra mitotic division. Conversely, mutations in Tor result in premature meiotic entry before the completion of the four mitotic divisions. Moreover, it was demonstrated that in the female germ line, the GATOR2 components Mio and Seh1 are required to oppose the TORC1 inhibitory activity of the Iml1/GATOR1 complex to prevent the constitutive down-regulation of TORC1 activity in later stages of oogenesis. These studies represent the first examination of the regulatory relationship between Iml1/GATOR1 and GATOR2 components within the context of a multicellular animal. Finally, these data reveal a surprising tissue-specific requirement for the GATOR2 complex in multicellular organisms and suggest a conserved role for the SEA/GATOR complex in the regulation of TORC1 activity during gametogenesis (Wei, 2014b).
Previous work demonstrated that in Drosophila the Iml1/GATOR1 complex mediates an adaptive response to amino acid starvation. This study tested the hypothesis that the Iml1/GATOR1 complex also has retained a role in the regulation of the early events of gametogenesis. Consistent with this model, this study found that in germline knockdowns of iml1, ovarian cysts delay meiotic entry and undergo a fifth mitotic division. This meiotic delay can be suppressed with the TORC1 inhibitor rapamycin. Thus, during Drosophila oogenesis the Iml1/GATOR1 complex promotes the transition from the mitotic cycle to the meiotic cycle through the down-regulation of the metabolic regulator TORC1. Increasing TORC1 activity by disabling its inhibitor delays meiotic progression, whereas germline clones of a Tor-null allele enter meiosis prematurely. Taken together, these data indicate that the level of TORC1 activity contributes to the timing of the mitotic/meiotic switch in Drosophila females and suggest that low TORC1 activity may be a conserved feature of early meiosis in many eukaryotes (Wei, 2014b).
However, in Drosophila, meiotic entry is not contingent on amino acid limitation at the organismal level. Indeed, the energy-intensive process of Drosophila oogenesis is curtailed dramatically when females do not have access to a protein source. Thus, to promote meiotic entry, Drosophila females must activate the Iml1/GATOR1 complex in a tissue-specific manner, using a mechanism that is independent of the overall nutrient status of the animal. At least two models can explain how Drosophila females might activate the Iml1/GATOR1 complex specifically in the germ line. In the first model, ovarian cysts locally experience low levels of amino acids during the mitotic cyst divisions and/or at the point of meiotic entry. These low levels of amino acids could reflect a non–cell-autonomous effect: The somatically derived escort cells that surround dividing ovarian cysts may function to create a low amino acid environment that triggers the activation of the Iml1/GATOR1 complex within developing ovarian cysts. Alternatively, the effect may be cell autonomous: The germ cells within dividing ovarian cysts may have a reduced ability to sense and/or import amino acids. In a second model, a developmental signaling pathway that is completely independent of local or whole-animal amino acid status directly activates the Iml1/GATOR1 complex. The identification of the upstream requirements for Iml1/GATOR1 activation in the female germ line will help distinguish between these two models (Wei, 2014b).
Although low TORC1 activity is required during early ovarian cyst development to promote the mitotic/meiotic switch, the dramatic growth of egg chambers later in oogenesis is a metabolically expensive process that is predicted to require high TORC1 activity. The current data indicate that the GATOR2 components Mio and Seh1 function to oppose the TORC1-inhibitory activity of the GATOR1 complex in the female germ line. In mio and seh1 mutants, TORC1 activity is constitutively repressed in the germ line of developing egg chambers, resulting in the activation of catabolic metabolism and the blocking of meiotic progression and oocyte development and growth (Wei, 2014b).
Previous data indicate that Mio and Seh1 act very early in oogenesis soon after the formation of the 16-cell cyst. The mio and seh1 ovarian phenotypes can be rescued by depleting the GATOR1 components nprl2, nprl3, or iml1 in the female germ line or by raising baseline levels of TORC1 activity by disabling an alternative TORC1 inhibitory complex, TSC1/2. These data are consistent with the model that the failure to maintain the meiotic cycle and the oocyte fate in mio and seh1 mutants is a direct result of inappropriately low TORC1 activity in the female germ line brought on by the deregulation of the Iml1/GATOR1 complex (Wei, 2014b).
Notably, null alleles of both mio and seh1 are viable, with many somatic tissues exhibiting no apparent developmental abnormalities and only limited reductions in cell growth. Thus, although Mio and Seh1 are critical for the activation of TORC1 and the development of the female gamete, these proteins play a relatively small role in the development and growth of many somatic tissues under nutrient-replete conditions. Whether this small role reflects the fact that components of the Iml1/GATOR1 complex are expressed at low levels in some somatic cell types or that the complex is present but needs to be activated by a signal, such as nutrient stress or a developmental signaling pathway, remains to be elucidated (Wei, 2014b).
In the future it will be important to gain a fuller understanding of the potential environmental and developmental inputs that regulate the activity of the Iml1/GATOR1 and GATOR2 complexes in multicellular organisms. These studies will provide much-needed insight into the basic mechanisms by which both environmental and developmental signaling pathways interface with the metabolic machinery to influence cell growth and differentiation (Wei, 2014b).
Target of Rapamycin Complex I (TORC1) is a central regulator of metabolism in eukaryotes that responds to a wide array of negative and positive inputs. The GTPase-activating protein toward Rags (GATOR) signaling pathway acts upstream of TORC1 and is comprised of two subcomplexes. The trimeric GATOR1 complex inhibits TORC1 activity in response to amino acid limitation by serving as a GTPase-activating protein (GAP) for the TORC1 activator RagA/B, a component of the lysosomally located Rag GTPase. The multi-protein GATOR2 complex inhibits the activity of GATOR1 and thus promotes TORC1 activation. This study reports that Wdr59, originally assigned to the GATOR2 complex based on studies performed in tissue culture cells, unexpectedly has a dual function in TORC1 regulation in Drosophila. ovary and the eye imaginal disc brain complex, Wdr59 inhibits TORC1 activity by opposing the GATOR2-dependent inhibition of GATOR1. Conversely, in the Drosophila fat body, Wdr59 promotes the accumulation of the GATOR2 component Mio and is required for TORC1 activation. Similarly, in mammalian HeLa cells, Wdr59 prevents the proteolytic destruction of GATOR2 proteins Mio and Wdr24. Consistent with the reduced levels of the TORC1-activating GATOR2 complex, Wdr59KOs HeLa cells have reduced TORC1 activity which is restored along with GATOR2 protein levels upon proteasome inhibition. Taken together, these data support the model that the Wdr59 component of the GATOR2 complex functions to promote or inhibit TORC1 activity depending on cellular context (Zhang, 2023).
The highly conserved Target of Rapamycin Complex 1 (TORC1) is a central regulator of growth and metabolism in eukaryotes. In the presence of positive upstream inputs, TORC1 promotes anabolic metabolism and growth by phosphorylating numerous downstream targets including S6K and 4EBP. Conversely, when exposed to negative cues, such as limited nutrients, or the lack of growth factors, cells downregulate TORC1 to activate catabolic metabolism and inhibit growth. The deregulation of TORC1 is implicated in a wide array of diseases including cancer, epilepsy, and aging. Thus, there is intense interest in obtaining a mechanistic understanding of how upstream signaling pathways regulate TORC1 activity (Zhang, 2023).
The Rag GTPase is a heterodimer comprised of a Rag A/B subunit and a RagC/D subunit, that recruits TORC1 to lysosomes for activation by the small GTPase Rheb, when the RagA/B component of the GTPase is in the GTP-bound state. The GTPase-activating protein toward Rags (GATOR) complex is an important upstream regulator of TORC1 that responds to the presence of nutrients. The GATOR complex, originally identified in yeast and named the Seh1 Associated (SEA) complex, is comprised of two subcomplexes, GATOR1 and GATOR2. The GATOR1 complex, which contains the proteins Nprl2, Nprl3, and DEPDC5/Iml1, inhibits TORC1 activity by serving as a GAP (GTPase-activating protein) for the lysosomally located Rag GTPase (Zhang, 2023).
The GATOR2 complex inhibits GATOR1 and thus serves to activate TORC1. However, the mechanism by which GATOR2 inhibits GATOR1 remains unknown. In its initial functional characterization in mammalian and Drosophila cultured cells, the GATOR2 complex was reported to contain five protein Mios/Mio, Seh1, Sec13, Wdr24, and Wdr59. In these studies, knockdowns of GATOR2 components resulted in the constitutive activation of GATOR1 and decreased TORC1 activity. Similarly, Drosophila mutants of the GATOR2 components Mio, Seh1, and Wdr24 exhibit decreased TORC1 activity and growth in the female germline. However, a recent study from Schizosaccharomyces pombe reported that SEA3/WDR59 inhibits TORC1 activity as a component of the GATOR1 complex. Notably, this is oppositive to the role assigned to Wdr59 based on studies in both human and Drosophila cultured cells. Additionally, deletions of Wdr59 in HEK293 cells and mouse embryonic fibroblasts result in a partial resistance to nutrient deprivation. Thus, the exact function of Wdr59 within the GATOR-TORC1 signaling pathway remains unclear (Zhang, 2023).
This study defines the in vivo requirement for the GATOR component Wdr59 in Drosophila. Wdr59 displays two distinct functions depending on cell type. First, in the ovary and the eye imaginal disc brain complex, Wdr59 acts upstream of the GATOR2 complex to inhibit TORC1 activity. Second, in the adult fat body and mammalian HeLa cells, Wdr59 protects the GATOR2 complex from proteolysis and promotes TORC1 activity. These data provide mechanistic insight into the complex role of the GATOR component Wdr59 in the tissue-specific regulation of TORC1 activity (Zhang, 2023).
The GATOR complex is an essential upstream regulator of TORC1 that is frequently mutated in human disease. This study describes a dual function for the GATOR component Wdr59 in TORC1 regulation in Drosophila. Surprisingly, Wdr59 functions were found to inhibit or promote TORC1 activity depending on cellular context. These studies broaden understanding of the GATOR-TORC1 signaling axis in metazoans and demonstrate the importance of examining metabolic regulation in vivo (Zhang, 2023).
In vivo studies often reveal tissue-specific and/or metabolic requirements for genes that are not observed in cell culture. In cultured cells from both mammals and Drosophila, RNAi depletions of wdr59 result in decreased TORC1 activity and slow growth, thus phenocopying depletions/knockouts of the GATOR2 components wdr24, seh1, and mio. Based on these initial studies, Wdr59 was assigned to the GATOR2 complex, which promotes TORC1 activity by downregulating the TORC1 inhibitor GATOR1. However, in vivo studies indicate wdr59 is not obligate member of the GATOR2 complex. Using Drosophila as a model, this study found that wdr59 mutant ovaries have increased TORC1 activity and growth rates and an attenuated response to nutrient stress. Notably, these phenotypes are the opposite of those reported for mutants of the GATOR2 components mio, seh1, and wdr24, but phenocopy those observed in mutants of the GATOR1 components nprl2, nprl3, and iml1. Thus, in the Drosophila ovary, Wdr59 functions to restrict TORC1 activity. In line with these findings, the binding of RagAGTP to the Nprl2 component of the GATOR1 complex was demonstrated to be decreased in wdr59 mutants, again the opposite of what was observed in mutants of the GATOR2 component wdr24. Taken together, these data suggest that wdr59 inhibits TORC1 activity by increasing the activity of the TORC1 inhibitor GATOR1. Notably, recent evidence from s. pombe proposes that Wdr59, known as SEA3 in yeast, acts to inhibit TORC1 as a component of the GATOR1 complex. However, epistasis analysis indicated that in Drosophila, Wdr59 is not a component of the GATOR1 complex but instead functions as an inhibitor of GATOR2 (Zhang, 2023).
How might wdr59 impact TORC1 activity? Biochemical, structural, and computational analysis from yeast indicate that SEA3/wdr59 is well positioned to function as an interacting hub connecting GATOR2 with GATOR1. Consistent with this observation, this study found that, as is observed in fission yeast, wdr59 regulates the interaction between components of the GATOR1 and GATOR2 complexes, with an increased association of GATOR2 with GATOR1 observed in the wdr59 mutant background. Thus, one possible model to explain these data is that wdr59 inhibits TORC1 activity by restricting the interaction of the GATOR2 complex with GATOR1, thus unleashing its GAP activity and TORC1 inhibitory potential. A prediction from this model is that components of the GATOR2 complex will be epistatic to Wdr59. In other words, Wdr59 requires the presence of an active GATOR2 complex to regulate TORC1 activity. Consistent with this prediction, it was fond that in the Drosophila ovary, wdr59, GATOR2 double mutants have a GATOR2-like phenotypes with reduced growth and TORC1 activity, strongly suggesting that GATOR2 is downstream of Wdr59. In contrast, in mutants or depletions of the GATOR1 components nprl2, nprl3, and Iml1, RagA remains in its TORC1-activating GTP-bound state, promoting the constitutive recruitment and activation of TORC1 on lysosomes independent of the status of GATOR2. Thus, GATOR1 components, unlike Wdr59, are epistatic to upstream members of the pathway including components of the GATOR2 complex. Taken together these data support the model that in Drosophila, Wdr59 is not required for GATOR1 function but instead acts upstream of the complex to regulate the activity of the GATOR1 inhibitor GATOR2. Importantly, the data demonstrate that the GATOR2 complex can inhibit GATOR1 independent of the Wdr59 subunit (Zhang, 2023).
Currently, there are conflicting reports on the role of Wdr59/Sea3 in the regulation of TORC1 activity. In Drosophila and mammalian cultured cells, as well as in mouse breast cancer tumors, Wdr59 promotes TORC1 activity while in fission yeast Wdr59 inhibits TORC1 activity. The current results provide a potential explanation for this contradiction. In Drosophila, Wdr59 either promotes or inhibits TORC1 activity depending on cell type. In the female germline and the eye imaginal disc brain complex, Wdr59 inhibits TORC1 activity. However, in the adult fat body, Wdr59 promotes TORC1 activity. Importantly, in the adult fat body, but not the ovary, Wdr59 is required for the accumulation of the Mio protein. Thus, a possible reason that wdr59 mutant fat bodies have reduced TORC1 activity is that they do not have a functional GATOR2 complex. This model is consistent with the finding that the GATOR2 complex is downstream of Wdr59 (Zhang, 2023).
To further explore why Wdr59 promotes TORC1 activation in some cellular contexts, the levels of GATOR2 proteins were examined in Wdr59KO HeLa cells. It was wondered if the TORC1 promoting function for Wdr59 observed in cultured cells might reflect the requirement for Wdr59 protein to accumulate components of the GATOR2 complex. Indeed, dramatically lower levels of the GATOR2 components Mios and Wdr24 were found in Wdr59KO HeLa cells due to proteolytic destruction by the proteosome. Strikingly, inhibiting the proteasome pharmacologically resulted in increased levels of GATOR2 proteins which was accompanied by a partial rescue of TORC1 activity. These data strongly suggest that the low TORC1 activity observed in Wdr59KD or Wdr59KO HeLa cells is due at least in part to the concomitant decrease in GATOR2 protein levels (Zhang, 2023).
Taken together, these data support the model that Wdr59 either promotes or inhibits TORC1 activity depending on cellular context. The GATOR-TORC1 signaling pathway is frequently cited as a potential target of pharmaceutical intervention because of its role in cancer and epilepsy. Thus, it is essential to have a full mechanistic understanding of the in vivo function of the GATOR complex in the regulation of TORC1 signaling and growth in metazoans (Zhang, 2023).
Prior to submission of this manuscript, a detailed cryo-electron microscopy structure of the human GATOR2 complex was published. It was reported that GATOR2 is a large 1.1 Mda complex that forms a cage-like structure, built on a continuous scaffold. As previously described, GATOR2 complex components contain numerous features common to membrane coating complexes which can form scaffolds that alter the curvature of membranes. Consistent with these studies, the authors report that the two copies of the Wdr59 subunit mediate the association of GATOR2 with GATOR1. However, as discussed above, it was found that Wdr59 opposes the association of GATOR2 with GATOR1 in several Drosophila tissues, the opposite of what is reported in this study in HEK293T cells. Whole animal studies in Drosophila have determined that there are unique tissue-specific requirements for multiple individual GATOR2 subunits, including Mio, Seh1, Wdr24, and now Wdr59. Going forward, it will be fascinating to determine how GATOR2 structure mediates tissue-specific GATOR2 functions in vivo (Zhang, 2023).
The nuclear pore complex (NPC) mediates the transport of macromolecules between the nucleus and cytoplasm. Recent evidence indicates that structural nucleoporins, the building blocks of the NPC, have a variety of unanticipated cellular functions. This study reports an unexpected tissue-specific requirement for the structural nucleoporin Seh1 during Drosophila oogenesis. Seh1 is a component of the Nup107-160 complex, the major structural subcomplex of the NPC. Seh1 associates with the product of the missing oocyte (mio) gene. In Drosophila, mio regulates nuclear architecture and meiotic progression in early ovarian cysts. Like mio, seh1 has a crucial germline function during oogenesis. In both mio and seh1 mutant ovaries, a fraction of oocytes fail to maintain the meiotic cycle and develop as pseudo-nurse cells. Moreover, the accumulation of Mio protein is greatly diminished in the seh1 mutant background. Surprisingly, characterization of a seh1 null allele indicates that, although required in the female germline, seh1 is dispensable for the development of somatic tissues. This work represents the first examination of seh1 function within the context of a multicellular organism. Seh1 has been shown to play a role in the construction and/or maintenance of bipolar spindles in multiple organisms. In summary, these studies demonstrate that Mio is a novel interacting partner of the conserved nucleoporin Seh1 and add to the growing body of evidence that structural nucleoporins can have novel tissue-specific roles (Senger, 2011).
The pathways that control progression through the early meiotic cycle remain poorly understood in metazoans. Drosophila melanogaster provides a genetically tractable system with which to study the relationship between early meiotic progression and oocyte development. As in mammals and Xenopus, the Drosophila oocyte initiates meiosis within the context of a germline cyst. Drosophila ovarian cysts are produced through a series of four synchronous mitotic divisions during which cytokinesis is incomplete. Soon after the completion of the mitotic divisions, all 16 cells enter premeiotic S phase. However, only the true oocyte, which comprises one of the two cells at the center of the syncytium, remains in meiosis and goes on to produce a gamete. The other 15 cells lose their meiotic features, enter the endocycle, and develop as polyploid nurse cells. In contrast to the nurse cells, the single oocyte remains in prophase of meiosis I until it proceeds to the first meiotic metaphase late in oogenesis. The pathways that drive this complicated series of cell cycle transitions that are so critical to the development of the mature gamete remain a topic of great interest (Senger, 2011).
The missing oocyte (mio) gene was identified in a forward genetic screen for mutants affecting cell cycle regulation and oocyte differentiation in early ovarian cysts (Iida, 2004). In mio mutants, the oocyte enters the meiotic cycle, forms mature synaptonemal complexes and accumulates oocyte-specific markers. However, in the absence of Mio, the oocyte fate is not stably maintained. Soon after the nurse cells enter the endocycle in stage 1 of oogenesis, mio oocytes follow the nurse cells into the endocycle, lose the preferential accumulation of oocyte-specific markers and develop as pseudo-nurse cells. Thus, mio is required for the maintenance of the meiotic cycle and oocyte identity. The mio gene encodes a 975 amino acid protein that is highly conserved from yeast to humans (Iida, 2004). Yet, the molecular function of mio remains elusive. This study demonstrates that Mio associates with the conserved nucleoporin Seh1 (also known as Nup44A in Drosophila). Moreover, this study define a tissue-specific requirement for Seh1 during oogenesis (Senger, 2011).
Seh1 is a component of a nucleoporin subcomplex known as the Nup107-160 complex in higher eukaryotes and the Nup84 complex in yeast (Fahrenkrog, 2004; Hetzer, 2005; Wozniak, 2010). The Nup107-160 complex, which is the major structural component of the nuclear pore complex (NPC), consists of at least nine subunits in higher eukaryotes and functions in the regulation of mRNA export as well as in the assembly and distribution of NPCs within the nuclear envelope (Hetzer, 2005; Wozniak, 2010). Studies over the last five years have defined several physiological functions for the Nup107-160/Nup84 complex that appear to be independent of nucleocytoplasmic transport (Fahrenkrog, 2004; Wozniak, 2010). Most notably, in Xenopus egg extracts and HeLa cells, the Nup107-160 complex has a dynamic localization during the cell cycle (Hetzer, 2005). Although present on the nuclear envelope in interphase, the entire complex targets to kinetochores, spindles and spindle poles to varying extents during mitosis (Loiodice, 2004; Orjalo, 2006). Consistent with a mitotic function, depleting components of the Nup107-160 complex results in cell cycle abnormalities, including defects in mitotic spindle formation, chromosome segregation and cytokinesis (Orjalo, 2006; Platani, 2009). Moreover, recent evidence indicates that in HeLa cells and Xenopus egg extracts, the Nup107-160 complex mediates microtubule nucleation at kinetochores via its interaction with the γ-TuRC complex (Mishra, 2010). Unlike in other metazoans, in Drosophila Nup107 fails to localize to kinetochores at mitosis but is found concentrated in the spindle region (Katsani, 2008). In summary, the Nup107-160 complex is multifunctional, with roles in both nucleocytoplasmic transport and cell cycle regulation (Senger, 2011).
This study demonstrates that Mio, a protein that is required for maintenance of the meiotic cycle and oocyte fate during oogenesis, associates with the structural nucleoporin Seh1. Surprisingly, it was found that a seh1 deletion allele is viable but exhibits dramatically reduced female fertility. Closer examination reveals that, as is observed in mio mutants, in a fraction of seh1 ovarian cysts oocytes fail to maintain the meiotic cycle and oocyte fate into later stages of oogenesis. From these studies it is concluded that Seh1 has an essential germline function during oogenesis but is not required for the growth or development of somatic tissues (Senger, 2011).
To better understand how Mio influences the maintenance of the meiotic cycle and oocyte fate in Drosophila, proteins that co-purify with Mio were identified by tandem affinity purification. From these experiments, it was determined that Mio is present in a stable complex with the structural nucleoporin Seh1. Seh1 is a component of the Nup107 complex, which is the primary structural unit of the nuclear pore. Studies in multiple organisms indicate that, although Seh1 is a nucleoporin, it is not required for bulk nucleocytoplasmic transport and has a limited role in the localization of other nucleoporins to the NPC (Loiodice, 2004; Orjalo, 2006). In contrast to its limited role at the NPC during interphase, recent evidence indicates that Seh1 has an essential function during mitosis. In Xenopus egg extracts and mammalian tissue culture cells, a fraction of the Nup107 complex that includes Seh1 targets to kinetochores, spindles and/or spindle poles from early prometaphase through early anaphase (Belgareh, 2001; Orjalo, 2006). Moreover, reducing the levels of Nup107-160 components disrupts spindle assembly and cytokinesis (Aitchison, 1995; Bai, 2004; Mishra, 2010; Orjalo, 2006; Platani, 2009; Zuccolo, 2007). Importantly, the specific depletion of seh1 results in the failure of the Nup107 complex to target to kinetochores at mitosis and results in multiple mitotic defects (Zuccolo, 2007; Platani, 2009). Thus, Seh1 plays a role in the construction and/or maintenance of bipolar spindles in multiple organisms (Senger, 2011).
To define the role of Seh1 in Drosophila a seh1null deletion allele was generated. Considering the key role of Seh1 during mitosis in other organisms, it was surprising to find that seh1null homozygotes are viable. From this observation it is concluded that seh1 is dispensable in Drosophila for the mitotic cycle during the majority of somatic divisions. Consistent with a limited role for Seh1 during the mitotic cycle, the specific accumulation of Seh1 on kinetochores during mitosis was not observed. Although this might reflect a limitation of the reagents, these data are in agreement with previous work (Katsani, 2008) demonstrating that the core component of the Nup107-160 complex, the nucleoporin Nup107, fails to accumulate at kinetochores during mitosis in multiple Drosophila tissues (Senger, 2011).
These studies also suggest a limited and/or redundant role for Seh1 in supporting the general structure and/or function of the NPC in interphase cells. Specifically, it was found that multiple nucleoporins, including Nup107, target to the NPC in the absence of Seh1 in both germline and somatic tissues. The only nucleoporin mislocalization observed in seh1null mutants involved a limited displacement of Mtor from the nuclear envelope to the nucleoplasm in meiotic cysts in the germarium. Mtor is a component of the nuclear basket and is present on the cytoplasmic face of the NPC during interphase but relocates to the spindle matrix during mitosis (Lince-Faria, 2009; Qi, 2004). Whether the partial displacement of Mtor in the seh1 background reflects a direct role for Seh1 in recruiting and/or stabilizing Mtor at the NPC remains to be determined (Senger, 2011).
Although dispensable for somatic development, it was found that seh1 has an essential function in the female germline during oogenesis. seh1null females are nearly sterile, producing only a small number of adult progeny. Indeed, seh1null females lay fewer eggs and contain ovarioles with a diminished number of egg chambers relative to wild-type females. A possible contributory factor to this reduced egg production is the mitotic delay observed during the ovarian cyst divisions. It was found that in ovaries from seh1 females, stem cells and ovarian cysts in R1 of the germarium spend a greater proportion of their time in mitosis than those in wild-type females. This phenotype is consistent with the metaphase delay observed in mammalian cells and Xenopus egg extracts depleted of members of the Nup107-160 complex, including Seh1. Thus, mutations in seh1 alter the rate of egg chamber production, as well as the nature of the ovarian cyst divisions in the germarium (Senger, 2011).
In addition to affecting the overall rate of egg production, Seh1 influences the differentiation of the oocyte within the ovarian cyst. Similar to what is observed in mio mutants, in a fraction of seh1 ovarian cysts the oocyte enters the endocycle and develops as a pseudo-nurse cell. This does not reflect an inability ofseh1null oocytes to enter the meiotic cycle. On the contrary, seh1null ovarian cysts enter the meiotic cycle on schedule with the two pro-oocytes progressing to pachytene, as measured by the construction of a mature synaptonemal complex. However, soon after exiting the germarium, a fraction of seh1 mutant oocytes enter the endocycle and become polyploid. In Drosophila, oocyte differentiation, as well as the maintenance of the meiotic cycle, are contingent on the microtubule-based transport of mRNAs and proteins from the nurse cells to the oocyte (Huynh, 2004). The germline-specific RNA-binding protein Orb starts to accumulate in the oocyte in late R2a of the germarium. Defects that impair the microtubule-dependent accumulation of Orb in the oocyte correlate with the inability to maintain the meiotic cycle through later stages of oogenesis. This study found that in seh1 ovarian cysts, the specific accumulation of Orb in the oocyte, as well as the secondary migration of Orb protein from the anterior to the posterior of the oocyte, are often delayed and/or otherwise defective. Additionally, the microtubule-dependent translocation of centrioles from the anterior to the posterior of the oocyte nucleus in the stage 1 oocyte is defective in both mio and seh1 ovarian cysts. Thus, as is observed with mio, seh1 influences the ability of the oocyte to maintain the meiotic cycle and oocyte fate beyond the germarium (Senger, 2011).
As Seh1 has been implicated in a variety of cellular functions, there are several possibilities as to how it might influence oocyte development and meiotic progression. First, Seh1 might act at the NPC to regulate the nucleocytoplasmic transport of specific molecules required for oocyte differentiation and growth. Second, Seh1 might regulate the activity of Mtor and/or other nucleoporins that have recently been implicated in transcriptional regulation (Vaquerizas, 2010). Finally, consistent with the alterations in mitotic cyst division and Orb localization, Seh1 might directly influence the organization and/or function of microtubules within ovarian cysts. Currently, the third model is favored because it is the most congruent with previous observations on the role of Seh1 in other organisms as well as with the current data (Senger, 2011).
This study has shown that Mio and Seh1 are present in a stable complex and that both proteins are dispensable for somatic development but are required for the development of the mature egg. Additionally, it was found that Seh1 is required for Mio protein stability. In the seh1 mutant background, Mio protein levels are reduced dramatically. Furthermore, depleting seh1 via RNAi in S2 tissue culture cells results in a rapid reduction in Mio protein levels. These results suggest the following simple model. Seh1 influences oocyte growth and the maintenance of the oocyte fate through its ability to promote the stability of the Mio protein. In the absence of Seh1, Mio protein levels fall, resulting in a mio-like phenotype. However, two lines of evidence suggest that Mio and Seh1 have a more complex interaction. First, overexpressing mio in the seh1 mutant background fails to rescue the seh1 phenotype. This failure to rescue is observed even though the seh1 15; UAS-mio ovaries have high levels of Mio protein in the germline. This strongly suggests that the seh1 ovarian phenotype is not due solely to the instability of the Mio protein in the absence of Seh1. Second, seh1 acts as a strong dominant suppressor of the mio 16-nurse-cell phenotype. A possible model to explain this counterintuitive result is that mio and seh1 act in opposing directions to regulate a common pathway that is crucial for the maintenance of the oocyte fate. Misregulation of this common pathway by either mio or seh1 could result in the reversion of the oocyte to the default state of nurse cell (Senger, 2011).
In the future, studies of Mio and Seh1 will help elucidate the pathways that drive oocyte development and meiotic progression and contribute to understanding of how individual NPC components drive tissue-specific differentiation (Senger, 2011).
TORC1 (see Drosophila Tor) is a master regulator of metabolism in eukaryotes that responds to multiple upstream signaling pathways. The GATOR complex is a newly defined upstream regulator of TORC1 that contains two sub-complexes, GATOR1, which inhibits TORC1 activity in response to amino acid starvation and GATOR2, which opposes the activity of GATOR1. The genome of Drosophila contains a single Sea2/Wdr24 homolog encoded by the gene CG7609 that shares 25% identity and 44% similarity to yeast Sea2 and 37% identity and 54% similarity to the human homolog WDR24. This study defines the in vivo role of the GATOR2 component Wdr24 in Drosophila. Wdr24 was shown to have both TORC1 dependent and independent functions in the regulation of cellular metabolism. Through the characterization of a null allele, it was shown that Wdr24 is a critical effector of the GATOR2 complex that promotes the robust activation of TORC1 and cellular growth in a broad array of Drosophila tissues. Additionally, epistasis analysis between wdr24 and genes that encode components of the GATOR1 complex revealed that Wdr24 has a second critical function, the TORC1 independent regulation of lysosome dynamics and autophagic flux. Notably, it was found that two additional members of the GATOR2 complex, Mio and Seh1, also have a TORC1 independent role in the regulation of lysosome function. Wdr24 was also shown to promotes lysosome acidification and autophagic flux in mammalian cells. Taken together these data support the model that Wdr24 is a key effector of the GATOR2 complex, required for both TORC1 activation and the TORC1 independent regulation of lysosomes (Cai, 2016).
In metazoans multiple conserved signaling pathways control the integration of metabolic and developmental processes. TORC1 is an evolutionarily conserved multi-protein complex that regulates metabolism and cell growth in response to an array of upstream inputs including nutrient availability, growth factors and intracellular energy levels. The catalytic component of TORC1 is the serine/threonine kinase Target of Rapamycin (TOR). When nutrients are abundant, TORC1 activity promotes translation, ribosome biogenesis as well as other pathways associated with anabolic metabolism and cell growth. However, when nutrients or other upstream activators are limiting, TORC1 activity is inhibited triggering catabolic metabolism and autophagy (Cai, 2016).
The Seh1 associated/GTPase-activating protein toward Rags (SEA/GATOR) complex is a newly identified upstream regulator of TORC1 that can be divided into two putative sub-complexes GATOR1 and GATOR2 (Bar-Peled, 2013; Dokudovskaya, 2011; Panchaud, 2013). The GATOR1 complex, known as the Iml1 complex or the Seh1 Associated Complex Inhibits TORC1 (SEACIT) in yeast, inhibits TORC1 activity in response to amino acid limitation. SEACIT/GATOR1 contains three proteins Npr2/Nprl2, Npr3/Nprl3 and Iml1/DEPDC5. Recent evidence, from yeast and mammals, indicates that the components of the SEACIT/GATOR1 complex function through the Rag GTPases to inhibit TORC1 activity. Notably, Nprl2 and DEPDC5 are tumor suppressor genes while mutations in DEPDC5 are a leading cause of hereditary focal epilepsies (Cai, 2016).
The GATOR2 complex, which is referred to as Sevh1 Associated Complex Activates TORC1 (SEACAT) in yeast, activates TORC1 by opposing the activity of GATOR1. The SEACAT/GATOR2 complex is comprised of five proteins, Seh1, Sec13, Sea4/Mio, Sea2/WDR24, and Sea3/WDR59. Computational analysis indicates that multiple components of the GATOR2 complex have structural features characteristic of coatomer proteins and membrane tethering complexes. In line with the structural similarity to proteins that influence membrane dynamics, in Drosophila the GATOR2 subunits Mio and Seh1 localize to multiple endomembrane compartments including lysosomes, the site of TORC1 regulation, and autolysosomes. In metazoans, members of the Sestrin and Castor family of proteins bind to and inhibit the GATOR2 complex in response to leucine and arginine starvation respectively. This interaction is proposed to inhibit TORC1 activity through the derepression of the GATOR1 complex. However, how GATOR2 opposes GATOR1 activity, thus allowing for the robust activation of TORC1, remains unknown. Additionally, the role of the GATOR2 complex in the regulation of both the development and physiology of multicellular animals remains poorly defined (Cai, 2016).
Recent evidence from Drosophila indicates that the requirement for the GATOR2 complex may be context specific in multicellular animals. In Drosophila, null alleles of the GATOR2 components mio and seh1 are viable but female sterile. Surprisingly, somatic tissues from mio and seh1 mutants exhibit little if any reductions in cell size and have nearly normal levels of TORC1 activity. In contrast, TORC1 activity is dramatically decreased in ovaries from mio and seh1 mutant females. This decrease in TORC1 activity is accompanied by the activation of catabolic metabolism in the female germ line, a dramatic reduction in egg chamber growth and difficulties maintaining the meiotic cycle. Thus, there is a surprising tissue specific requirement for the GATOR2 components Mio and Seh1 during oogenesis. However, the in vivo role of the other members of the GATOR2 complex in the regulation of cellular metabolism remains undefined (Cai, 2016).
This study defines the in vivo requirement for the GATOR2 component Wdr24 in Drosophila. Wdr24 was found to have two distinct functions. First, Wdr24 is a critical effector of the GATOR2 complex that promotes TORC1 activity and cellular growth in a broad array of tissues. Second, Wdr24 is required for the TORC1 independent regulation of lysosome function and autophagic flux. Notably, two additional members of the GATOR2 complex, Mio and Seh1, also have a TORC1 independent role in the regulation of lysosome function. Taken together these data support the model that multiple components of the GATOR2 complex have both TORC1 dependent and independent roles in the regulation of cellular metabolism (Cai, 2016).
This study describes a dual role for the GATOR2 component Wdr24 in the regulation of TORC1 activity and lysosome dynamics. Wdr24 is a critical effector of the GATOR2 complex that promotes TORC1 activity in both germline and somatic tissues. This lies in contrast to the GATOR2 components Mio and Seh1, which have a limited role in the regulation of TORC1 activity in many cell types. Surprisingly, a second function of Wdr24 was identified that is independent of TORC1 status, the regulation of lysosome acidification and autophagic flux. Taken together these data support the model that the GATOR2 complex regulates both the response to amino acid starvation and lysosome function (Cai, 2016).
Whole animal studies often reveal tissue-specific and/or metabolic requirements for genes that are not readily observed in cell culture. In mammalian and Drosophila tissue culture cells, RNAi based depletions of the GATOR2 components Mio, Seh1, Wdr59, and Wdr24 result in decreased TORC1 activity in return to growth assays (Bar-Peled, 2013, Wei, 2014). These data have resulted in the model that all components of the GATOR2 complex are generally required for TORC1 activation (Wei, 2014). However, the characterization of mio and seh1 null mutants in Drosophila, demonstrated that Mio and Seh1 are critical for the activation of TORC1 and inhibition of autophagy in the female germ line, but play a relatively small role in the regulation of TORC1 activity and autophagy in somatic tissues under standard culture conditions. Thus, the requirement for at least a subset of GATOR2 complex components is tissue and/or context specific (Cai, 2016).
This study reports that the GATOR2 component Wdr24 is required for the full activation of TORC1 in both germline and somatic cells of Drosophila. Consistent with the global down-regulation of TORC1 activity in the absence of Wdr24, wdr24 mutant adults are notably smaller than controls and are female sterile. Depleting the GATOR1 components nprl2 and nprl3 in the wdr24 mutant background rescued the low TORC1 activity, growth defects, and female sterility of wdr24 mutants. Thus, the GATOR2 component Wdr24 is required to oppose GATOR1 activity in both germline and somatic cells of Drosophila. From these results it is proposed that Wdr24 is a key effector of the GATOR2 complex required for the full activation of TORC1 in most cell types (Cai, 2016).
There are several potential models to explain the differential requirement for individual GATOR2 proteins in Drosophila. First, there may be tissue specific requirements for individual GATOR2 subunits. In this model the different phenotypes observed in the seh1 and mio versus wdr24mutants reflects a qualitative difference in the requirement for these proteins in different tissues. However, an alternative model is favored in which Wdr24 is the core effector of GATOR2 activity, with Mio and Seh1 functioning primarily as positive regulators of GATOR2 activity. In this second model, the differential phenotypes observed in the seh1 and mio versus wdr24 mutants reflects a quantitative difference in the requirement for GATOR2 activity in different tissues. The distinction between these two models awaits the identification of the molecular mechanism of Wdr24 and GATOR2 action (Cai, 2016).
A novel TORC1 independent role has been identified for Wdr24 in the regulation of lysosome dynamics and function. In wdr24 mutants, the down-regulation of TORC1 activity and the accumulation of autolysosomes occur independent of nutrient status. It was initially hypothesized that in the absence of the GATOR2 component Wdr24, the deregulation of the GATOR1 complex results in low TORC1 activity, triggering the constitutive activation of autophagy and the accumulation of autolysosomes. Surprisingly, however, epistasis analysis determined that the accumulation of lysosomes could be decoupled from both the chronic inhibition of TORC1 activity and the activation of autophagy. Raising TORC1 activity in the wrd24 mutant background, by depleting either components of the GATOR1 or TSC complex, failed to rescue the accumulation of abnormal lysosomal structures. Notably, it was determined that two additional members of the GATOR2 complex, Mio and Seh1, also regulate lysosomal behavior independent of both GATOR1 and the down-regulation of TORC1 activity. From these data it is inferred that multiple components of the GATOR2 complex have a TORC1 independent role in the regulation of lysosomes (Cai, 2016).
An increased number of autolysosomes is often associated with reduced autophagic flux due to diminished lysosomal degradation. Consistent with reduced autophagic flux, in Drosophila wrd24-/- mutants accumulated enlarged autolysosomes filled with undegraded material. Moreover, lysosomes in the wrd24-/- mutants failed to quench the GFP fluorescence of a GFP-mCherry-Atg8a protein. These phenotypes are consistent with decreased lysosomal pH and degradative capacity. In order to examine in detail the role of Wdr24 in the regulation of lysosome function a wrd24-/- knockout HeLa cell line was generated that recapitulated the phenotypes observed in Drosophila wrd24-/- mutants. Specifically, wrd24-/- HeLa cells had have decreased TORC1 activity and accumulate a large number of autolysosomes. Using multiple assays it was determined that wrd24-/- lysosomes had reduced degradative capacity and autophagic flux and thus accumulate proteins that are normally degraded by lysosomal enzymes such as p62, LC3II and Cathepsin D. Additionally, it was determined that wrd24-/-lysosomes have increased pH relative to wild-type cells, again consistent with reduced lysosomal function. Taken together these data confirm that Wdr24 plays a key role in the regulation of lysosomal activity (Cai, 2016).
This study shows that components of the GATOR2 complex function in the regulation of TORC1 activity and in the TORC1 independent regulation of lysosomal dynamics and autophagic flux. These two functions suggest that the GATOR2 complex may regulate cellular homeostasis by coordinating TORC1 activity with the dynamic regulation of lysosomes during periods of nutrient stress. Intriguingly, several recent reports describe a very similar dual function for the RagA/B GTPases in both mice and zebrafish (Kim, 2014; Shen,2016). RagA/B play a critical role in the activation of TORC1 in the presence of amino acids (Kim, 2008; Sancak, 2008). Surprisingly, however, TORC1 activity was not found to be significantly decreased in cardiomyocytes of RagA/B knockout mice (Cai, 2016).
Nevertheless, the RagA/B mutant cardiomyocytes have decreased autophagic flux and reduced lysosome acidification. From published data, it was concluded that the RagA/B GTPases regulate lysosomal function independent of their role in the regulation of TORC1 activation in some cell types. Similarly, RagA is required for proper lysosome function and phagocytic flux in microglia. Notably, Mio, a component of the GATOR2 complex is found associated with RagA (Bar-Peled, 2013). Thus, in the future it will be important to determine if components of the GATOR2 complex function in a common pathway with the Rag GTPases to regulate lysosomal function (Cai, 2016).
In Saccharomyces cerevisiae single mutants of wrd24/sea2/ and wdr59/sea3 do not exhibit defects in TORC1 regulation but do have defects in vacuolar structure. Moreover, several recently identified genes that regulate the GATOR2-GATOR1-TORC1 pathway in response to amino acid limitation are restricted to metazoans. These data make it tempting to speculate that the ancestral function of the GATOR2 complex maybe the regulation of lysosome/vacuole function and autophagic flux. Indeed, the finding that GATOR2 components regulate lysosome dynamics is particularly intriguing in light of the observation that GATOR2 complex is comprised of proteins with characteristics of coatomer proteins and membrane tethering complexes. Notably, the GATOR2 complex components Mio, Seh1 and Wdr24 localize to lysosomes and autolysosomes. Similarly, these proteins associate with the vacuolar membrane in budding yeast. Thus, going forward it will be important to examine if the GATOR2 complex acts directly on lysosomal membranes to regulate their structure and/or function. More broadly, future studies on the diverse roles of the SEACAT/GATOR2 complex will further understanding of the complex relationship between cellular metabolism and the regulation of endomembrane dynamics in both development and disease (Cai, 2016).
In Drosophila, a single oocyte develops within a 16-cell germline cyst. Although all 16 cells initiate meiosis and undergo premeiotic S phase, only the oocyte retains its meiotic chromosome configuration and remains in the meiotic cycle. The other 15 cells in the cyst enter the endocycle and develop as polyploid nurse cells. A longstanding goal in the field has been to identify factors that are concentrated or activated in the oocyte, that promote meiotic progression and/or the establishment of the oocyte identity. This study presents the characterization of the missing oocyte gene, an excellent candidate for a gene directly involved in the differentiation of the oocyte nucleus. The missing oocyte gene encodes a highly conserved protein that preferentially accumulates in pro-oocyte nuclei in early prophase of meiosis I. In missing oocyte mutants, the oocyte enters the endocycle and develops as a polyploid nurse cell. Genetic interaction studies indicate that missing oocyte influences meiotic progression prior to pachytene and may interact with pathways that control DNA metabolism. These data strongly suggest that the product of the missing oocyte gene acts in the oocyte nucleus to facilitate the execution of the unique cell cycle and developmental programs that produce the mature haploid gamete (Iida, 2004; PubMed).
The TORC regulator GATOR1/SEACIT controls meiotic entry and early meiotic events in yeast. However, how metabolic pathways influence meiotic progression in metazoans remains poorly understood. This study examined the role of the TORC regulators GATOR1 and GATOR in the response to meiotic double-stranded breaks (DSB) during Drosophila oogenesis. In mutants of the GATOR component mio, meiotic DSBs trigger the constitutive downregulation of TORC activity and a permanent arrest in oocyte growth. Conversely, in GATOR mutants, high TORC activity results in the delayed repair of meiotic DSBs and the hyperactivation of p53. Unexpectedly, it was found that GATOR inhibits retrotransposon expression in the presence of meiotic DSBs in a pathway that functions in parallel to p53. Thus, these studies have revealed a link between oocyte metabolism, the repair of meiotic DSBs and retrotransposon expression (Wei, 2019).
Metabolism impacts meiotic progression during oogenesis. Target of Rapamycin Complex 1 (TORC1) is a multi-protein complex that functions as a master regulator of metabolism. In the presence of adequate nutrients and positive upstream growth signals, TORC1, which contains the serine/threonine kinase Target of Rapamycin, becomes active and functions to stimulate growth and inhibit catabolic metabolism through the phosphorylation of down-stream effector proteins. The Seh1 Associated Complex Inhibits TORC1 (SEACIT), originally identified in yeast, inhibits TORC1 activity in response to amino acid limitation. SEACIT, known as the GAP Activity Towards Rags complex 1 (GATOR1) in metazoans, is comprised of three highly conserved proteins Npr2/Nprl2, Npr3/Nprl3 and Iml1/Depdc5. In Drosophila and mammals, depleting any of the three GATOR1 components results in increased TORC1 activity and growth, as well as a reduced response to amino acid starvation. Thus, the role of the SEACIT/GATOR1 complex in the regulation of TORC1 activity is highly conserved in eukaryotes (Wei, 2019).
The multi-protein GATOR2 complex, known as Seh1 Associated Complex Activates TORC1 (SEACAT) in yeast, inhibits the activity of GATOR1 and thus functions to activate TORC1 (see Mio prevents the constitutive downregulation of TORC1 activity in response to meiotic DSBs). In metazoans, the GATOR2 complex functions in multiple amino acid sensing pathways. In tissue culture cells, depleting GATOR2 components results in the constitutive activation of GATOR1 and the permanent downregulation of TORC1 activity. However, genetic studies of the role of individual GATOR2 components in Drosophila, indicate that the requirement for the GATOR2 complex is more nuanced when examined in the context of a multicellular animal. For example, mutations in the GATOR2 component mio, result in a block to oocyte growth and differentiation, due to the constitutive downregulation of TORC1 activity in the female germline. However, mio is not required to maintain TORC1 activity in most somatic tissues of Drosophila (Wei et al., 2016). Why there is a tissue specific requirement for mio in the female germline of Drosophila is currently unknown (Wei, 2019).
In single celled eukaryotes, nutrient limitation often facilitates meiotic entry. In the yeast Saccharomyces cerevisiae, the down-regulation of TORC1 by SEACIT/GATOR1 in response to amino acid stress promotes both meiotic entry and early meiotic progression. Surprisingly, as is observed in yeast, during Drosophila oogenesis the GATOR1 complex promotes meiotic entry. These data raise the intriguing possibility that in Drosophila the GATOR1 complex and low TORC1 activity may be critical to the regulation of additional events of the early meiotic cycle (Wei, 2019).
This study reports that the GATOR complex is critical to the response to meiotic DSB during Drosophila oogenesis. Restraining TORC1 activity via a pathway that involves both GATOR1 and the Tuberous sclerosis complex (TSC) promotes the timely repair of meiotic DSBs and prevents the hyperactivation of p53 in the female germline. Notably, the delayed repair of meiotic DSBs in GATOR1 mutants is due, at least in part, to the hyperactivation of the TORC1 target S6K. Conversely, the data indicate that the GATOR2 component Mio opposes the activity of GATOR1 in the female germline, thus preventing the constitutive downregulation of TORC1 activity and allowing for the growth and development of the oocyte in later stages of oogenesis. Thus, this study has identified a regulatory loop required to modulate TORC1 activity in response to meiotic DSBs during Drosophila oogenesis. Finally, during the course of these studies, it was observed that the GATOR1 complex prevents the derepression of retrotransposon expression in the presence of meiotic DSBs (Wei, 2019).
Previous work has shown that in Drosophila, mutations in the GATOR2 component mio, result in the constitutive activation of the GATOR1 pathway in the female germline but not in somatic tissues. This study demonstrates that the tissue specific requirement for mio during oogenesis is due, at least in part, to the generation of meiotic DBSs during oogenesis. In Drosophila, only the female germline undergoes meiotic recombination and thus experiences the genotoxic stress associated with developmentally programmed DSBs. This study shows that in mio mutants, blocking the formation of meiotic DSBs prevents the constitutive downregulation of TORC1 activity thus allowing for the growth and development of the oocyte. These data are consistent with the model that meiotic DSBs trigger the activation of a TORC1 inhibitory pathway that must be opposed and/or attenuated by the GATOR2 component Mio (see A working model for the role of the GATOR complex in the response to meiotic DSBs) (Wei, 2019).
While there are several possible models that might explain these data, it is believed the most parsimonious explanation for these results is that the TORC1 inhibitory pathway activated by meiotic DSBs, involves both GATOR1 and TSC. This model is consistent with the ability of both GATOR1 and TSC depletions to rescue the mio mutant phenotype. Additionally, recent reports indicate that GATOR1 and TSC act in a common pathway to downregulate TORC1 activity in response to multiple upstream inhibitory inputs. Previously, it was determined that in Drosophila, amino acid starvation induces a dramatic GATOR1/TSC dependent decrease in TORC1 activity in somatic tissues, that far exceeds any reduction in TORC1 activity observed in GATOR2 null mutants. This observation strongly suggests that, in addition to the removal of the GATOR2 inhibition of GATOR1, there is an activation step that is required to fully potentiate the GATOR1/TSC pathway (Wei, 2019).
Thus, based on these data the following model is proposed. Meiotic DSBs activate, or are required to maintain, a GATOR1/TSC dependent pathway that downregulates TORC1 activity in the female germline. The GATOR2 component Mio is required to oppose or turnoff this pathway to prevent the constitutive downregulation of TORC1 activity in later stages of oogenesis. While it is believed that the data support the role of the GATOR1/TSC pathway, it is conceded that an alternative regulator of TORC1 activity may also be critical to the downregulation of TORC1 activity in response to meiotic DSBs (Wei, 2019).
Hyperactivation of TORC1 has been linked to defects in the DNA damage response in single celled and multicellular organisms. The observation that meiotic DSBs likely promote the GATOR1 dependent downregulation of TORC1 activity during Drosophila oogenesis, suggested that limiting TORC1 activity may be important to the regulation of meiotic DSB repair. In previous work, it was found that GATOR1 mutant ovaries had TORC1 activity levels approximately three times higher than those observed in wild-type ovaries. This study demonstrates that GATOR1 mutant ovaries exhibit multiple phenotypes consistent with the misregulation of meiotic DSB repair including, an increase in the steady state number of Mei-W68/Spo-11 induced DSBs, the retention of meiotic DSBs into later stages of oogenesis and the hyperactivation of p53. Importantly, RNAi depletions of Tsc1 partially phenocopied the GATOR1 ovarian defects. Thus, the misregulation of meiotic DSBs observed in GATOR1 mutant oocytes are due to high TORC1 activity and not to a TORC1 independent function of the GATOR1 complex (Wei, 2019).
Epistasis analysis between the GATOR1 component nprl3 and the Rad51 homolog spnA, strongly suggest that GATOR1 impacts the repair, rather than the generation, of meiotic DSBs. This study determined that double mutants of nprl2 and the Rad51 homolog spnA, which is required for the repair of meiotic DSBs, have approximately the same number of DSBs as spnA single mutants. These data are consistent with GATOR1 and spnA influencing the common process of DNA repair and are inconsistent with GATOR1 mutants producing supernumerary breaks (Wei, 2019).
These observations on the role of the GATOR1 complex during Drosophila oogenesis are particularly intriguing in light of similar meiotic defects observed in a npr3 mutants in Saccharomyces cerevisiae. In the sporulation proficient strain SK1, npr3 mutant cells enter meiosis and express the transcription factor and master regulator of gametogenesis IME1 with wild-type kinetics. Subsequently, npr3 mutants exhibit a mild delay in the generation of meiotic DNA breaks, but a substantial delay in the repair of meiotic DSBs. Thus, yeast and Drosophila SEACIT/GATOR1 mutants share a common meiotic phenotype, the delayed repair of meiotic DSBs. These results raise the intriguing possibility that low TORC1 activity may be a common feature of the early meiotic cycle in many organisms (Wei, 2019).
Notably, the data indicate that the delay in the repair of meiotic DSBs in GATOR1 mutants is due to the hyperactivation of the TORC1 downstream target S6K. S6K is a critical downstream effector of TORC1 that impacts multiple essential cellular processes including, but not limited to cell growth, energy balance and aging. Intriguingly, in mammals, S6K has been implicated in the regulation of the DNA damage response with hyperactivation of the TORC1-S6K pathway resulting in the accumulation of unrepaired DSBs and genome instability. Thus, similar to what is reported in mammals, the data are consistent with the model that the hyperactivity of the TORC1/S6K axis delays the repair of DSBs in Drosophila (Wei, 2019).
Finally, it was determined that GATOR1 mutants have a diminished response to DSBs outside the female germline in somatic tissues of Drosophila. Similar to what is observed in TSC mutant cells in humans that have increased levels of TORC1 activity, this study find that GATOR1 mutant embryos have a reduced ability to survive low levels of γ-irradiation. Moreover, in the somatic follicle cells of the ovary a delay was observed in the repair of DSBs after adult females are exposed to low levels of γ-irradiation. Thus, in Drosophila inappropriately high TORC1 activity delays the repair of DSBs in both the germline and somatic tissues (Wei, 2019).
The initiation of homologous recombination through the programmed generation of DNA double-stranded breaks (DSBs) is a universal feature of meiosis. DSBs represent a dangerous form of DNA damage that can result in dramatic and permanent changes to the germline genome. To minimize this destructive potential, the generation and repair of meiotic DSBs is tightly controlled in space and time . The activation of transposable elements represents an additional threat to genome integrity in germ line cells. Genotoxic stress, resulting from DNA damage, has been implicated in the deregulation of transposons in multiple organisms. Thus, germ line cells may be at an increased risk for transposon derepression due to the genotoxic stress associated with meiotic recombination. Consistent with this hypothesis, germ line cells have evolved extensive surveillance systems to detect and silence transposons beyond the pathways present in most somatic tissues (Wei, 2019).
Previous studies have shown that DNA damage promotes the deregulation of retrotransposon in multiple organisms, including Drosophila. In line with these studies, this study found that in GATOR1 mutants, the DSBs that initiate meiotic recombination trigger the deregulation of retrotransposon expression. Similarly, p53 mutant females derepress retrotransposon expression during oogenesis, but as observed in GATOR1 mutants, primarily in the presence of meiotic DSBs. Double mutants of nprl3, p53 exhibit a dramatic increase in retrotransposon expression relative to either p53 or nprl3 single mutants, implying that p53 and GATOR1 act through independent pathways to repress retrotransposon expression in the female germline. One possibility is that both GATOR1 and p53 independently impact genome stability. Thus, disabling both pathways may have an additive effect on both genome stability and retrotransposon expression. Consistent with the hypothesis that genome instability drives retrotransposon expression, this study found that mutants in spnA/Rad51, which fail to repair meiotic DSBs, also exhibit increased transcription of multiple retrotransposons. Intriguingly, the SpnA homolog Rad51, as well as other genes required for DNA repair, was recently identified in a high throughput screen for genes that suppress (Long Interspersed Element-1) LINE1 expression in mammalian tissue culture cells (Wei, 2019).
However, the current data also suggest that the GATOR1 complex may influence retrotransposon expression independent of the regulation of TORC1 activity. While both GATOR1 and TSC are required for the efficient repair of meiotic DSBs, in contrast to GATOR1 mutant ovaries, little to no increase was observed in retrotransposon expression in the Tsc1 depleted ovaries. We believe this reflects the incomplete depletion of Tsc1 by RNAi resulting in a reduced retention of meiotic DSBs relative to GATOR1 mutants. However, a second possibility is that the GATOR1 complex inhibits retrotransposon expression independent of TORC1 inhibition. As is observed with spnA the depletion of GATOR1 components, but not TSC components result in the activation of LINE1 expression in HeLa cells. Taken together, these data hint that the GATOR1 complex may impact retrotransposon expression in the germline via two independent pathways: First by promoting the repair of meiotic DSBs through the downregulation of TORC1 activity and second via a pathway that functions independent of TORC1 inhibition (Wei, 2019).
Genes encoding components of the GATOR1 complex are often deleted in cancers. As is observed in GATOR1 mutants, cancer cells frequently have increased TORC1 activity, increased genomic instability and increased retrotransposon expression. Thus, in the future it will be important to identify the molecular mechanism by which the GATOR1 complex influences both the response to genotoxic stress and the expression of retrotransposons under both normal and pathological conditions (Wei, 2019).
Search PubMed for articles about Drosophila Missing oocyte
Aitchison, J. D., Blobel, G., and Rout M. P. (1995). Nup120p: a yeast nucleoporin required for NPC distribution and mRNA transport. J. Cell Biol. 131: 1659-1675. PubMed ID: 8557736
Bai, S. W., et al. (2004). The fission yeast Nup107-120 complex functionally interacts with the small GTPase Ran/Spi1 and is required for mRNA export, nuclear pore distribution, and proper cell division. Mol. Cell. Biol. 24: 6379-6392. PubMed ID: 15226438
Bar-Peled, L., Chantranupong, L., Cherniack, A. D., Chen, W. W., Ottina, K. A., Grabiner, B. C., Spear, E. D., Carter, S. L., Meyerson, M. and Sabatini, D. M. (2013). A Tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science 340: 1100-1106. PubMed ID: 23723238
Belgareh, N., et al. (2001). An evolutionarily conserved NPC subcomplex, which redistributes in part to kinetochores in mammalian cells. J. Cell Biol. 154: 1147-1160. PubMed ID: 11564755
Cai, W., Wei, Y., Jarnik, M., Reich, J. and Lilly, M. A. (2016). The GATOR2 component Wdr24 regulates TORC1 activity and lysosome function. PLoS Genet 12: e1006036. PubMed ID: 27166823
Dokudovskaya, S., Waharte, F., Schlessinger, A., Pieper, U., Devos, D. P., Cristea, I. M., Williams, R., Salamero, J., Chait, B. T., Sali, A., Field, M. C., Rout, M. P. and Dargemont, C. (2011). A conserved coatomer-related complex containing Sec13 and Seh1 dynamically associates with the vacuole in Saccharomyces cerevisiae. Mol Cell Proteomics 10: M110 006478. PubMed ID: 21454883
Fahrenkrog, B., Koser, J. and Aebi, U. (2004). The nuclear pore complex: a jack of all trades? Trends Biochem. Sci. 29: 175-182. PubMed ID: 15082311
Hetzer, M. W., Walther, T. C. and Mattaj, I. W. (2005). Pushing the envelope: structure, function, and dynamics of the nuclear periphery. Annu. Rev. Cell Dev. Biol. 21: 347-380. PubMed ID: 16212499
Huynh, J. R. and St Johnston, D. (2004). The origin of asymmetry: early polarisation of the Drosophila germline cyst and oocyte. Curr Biol 14: R438-449. PubMed ID: 15182695
Iida, T. and Lilly, M. A. (2004). missing oocyte encodes a highly conserved nuclear protein required for the maintenance of the meiotic cycle and oocyte identity in Drosophila. Development 131: 1029-1039. PubMed ID: 14973288
Katsani, K. R., Karess, R. E., Dostatni, N. and Doye V. (2008). In vivo dynamics of Drosophila nuclear envelope components. Mol. Biol. Cell 19: 3652-3666. PubMed ID: 18562695
Kim, E., Goraksha-Hicks, P., Li, L., Neufeld, T. P. and Guan, K. L. (2008). Regulation of TORC1 by Rag GTPases in nutrient response. Nat Cell Biol 10: 935-945. PubMed ID: 18604198
Kim, Y. C., Park, H. W., Sciarretta, S., Mo, J. S., Jewell, J. L., Russell, R. C., Wu, X., Sadoshima, J. and Guan, K. L. (2014). Rag GTPases are cardioprotective by regulating lysosomal function. Nat Commun 5: 4241. PubMed ID: 24980141
Lince-Faria, M., et al. (2009). Spatiotemporal control of mitosis by the conserved spindle matrix protein Megator. J. Cell Biol. 184: 647-657. PubMed ID: 19273613
Loiodice, I., et al. (2004). The entire Nup107-160 complex, including three new members, is targeted as one entity to kinetochores in mitosis. Mol. Biol. Cell 15: 3333-3344. PubMed ID: 15146057
Mishra, R. K., et al. (2010). The Nup107-160 complex and gamma-TuRC regulate microtubule polymerization at kinetochores. Nat. Cell Biol. 12: 164-169. PubMed ID: 20081840
Orjalo, A. V., et al. (2006). The Nup107-160 nucleoporin complex is required for correct bipolar spindle assembly. Mol. Biol. Cell 17: 3806-3818. PubMed ID: 16807356
Panchaud, N., Peli-Gulli, M. P. and De Virgilio, C. (2013). Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Sci Signal 6: ra42. PubMed ID: 23716719
Platani, M., et al. (2009). The Nup107-160 nucleoporin complex promotes mitotic events via control of the localization state of the chromosome passenger complex. Mol. Biol. Cell 20, 5260-5275. PubMed ID: 16807356
Qi, H., et al. (2004). Megator, an essential coiled-coil protein that localizes to the putative spindle matrix during mitosis in Drosophila. Mol. Biol. Cell 15: 4854-4865. PubMed ID: 15356261
Sancak, Y., Peterson, T. R., Shaul, Y. D., Lindquist, R. A., Thoreen, C. C., Bar-Peled, L. and Sabatini, D. M. (2008). The Rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 320: 1496-1501. PubMed ID: 18497260
Senger, S., Csokmay, J., Akbar, T., Jones, T. I., Sengupta, P. and Lilly, M. A. (2011).The nucleoporin Seh1 forms a complex with Mio and serves an essential tissue-specific function in Drosophila oogenesis. Development 138(10): 2133-42. PubMed ID: 21521741
Shen, K., Sidik, H. and Talbot, W. S. (2016). The Rag-Ragulator complex regulates lysosome function and phagocytic flux in microglia. Cell Rep 14: 547-559. PubMed ID: 26774477
Vaquerizas, J. M., et al. (2010). Nuclear pore proteins nup153 and megator define transcriptionally active regions in the Drosophila genome. PLoS Genet. 6: e1000846. PubMed ID: 20174442
Wei, Y. and Lilly, M. A. (2014a). The TORC1 inhibitors Nprl2 and Nprl3 mediate an adaptive response to amino-acid starvation in Drosophila. Cell Death Differ 21: 1460-1468. PubMed ID: 24786828
Wei, Y., Reveal, B., Reich, J., Laursen, W. J., Senger, S., Akbar, T., Iida-Jones, T., Cai, W., Jarnik, M. and Lilly, M. A. (2014b). TORC1 regulators Iml1/GATOR1 and GATOR2 control meiotic entry and oocyte development in Drosophila. Proc Natl Acad Sci U S A 111(52):E5670-7. PubMed ID: 25512509
Wei, Y., Bettedi, L., Ting, C. Y., Kim, K., Zhang, Y., Cai, J. and Lilly, M. A. (2019). The GATOR complex regulates an essential response to meiotic double-stranded breaks in Drosophila. Elife 8. PubMed ID: 31650955
Wozniak, R., Burke, B. and Doye, V. (2010). Nuclear transport and the mitotic apparatus: an evolving relationship. Cell. Mol. Life Sci. 67: 2215-2230. PubMed ID: 20372967
Zhang, Y., Ting, C. Y., Yang, S., Reich, J., Fru, K. and Lilly, M. A. (2023). Wdr59 promotes or inhibits TORC1 activity depending on cellular context. Proc Natl Acad Sci U S A 120(1): e2212330120. PubMed ID: 36577058
Zuccolo, M., et al. (2007). The human Nup107-160 nuclear pore subcomplex contributes to proper kinetochore functions. EMBO J. 26: 1853-1864. PubMed ID: 17363900
date revised: 10 September 2025
Home page: The Interactive Fly © 2025 Thomas Brody, Ph.D.
Biological Overview
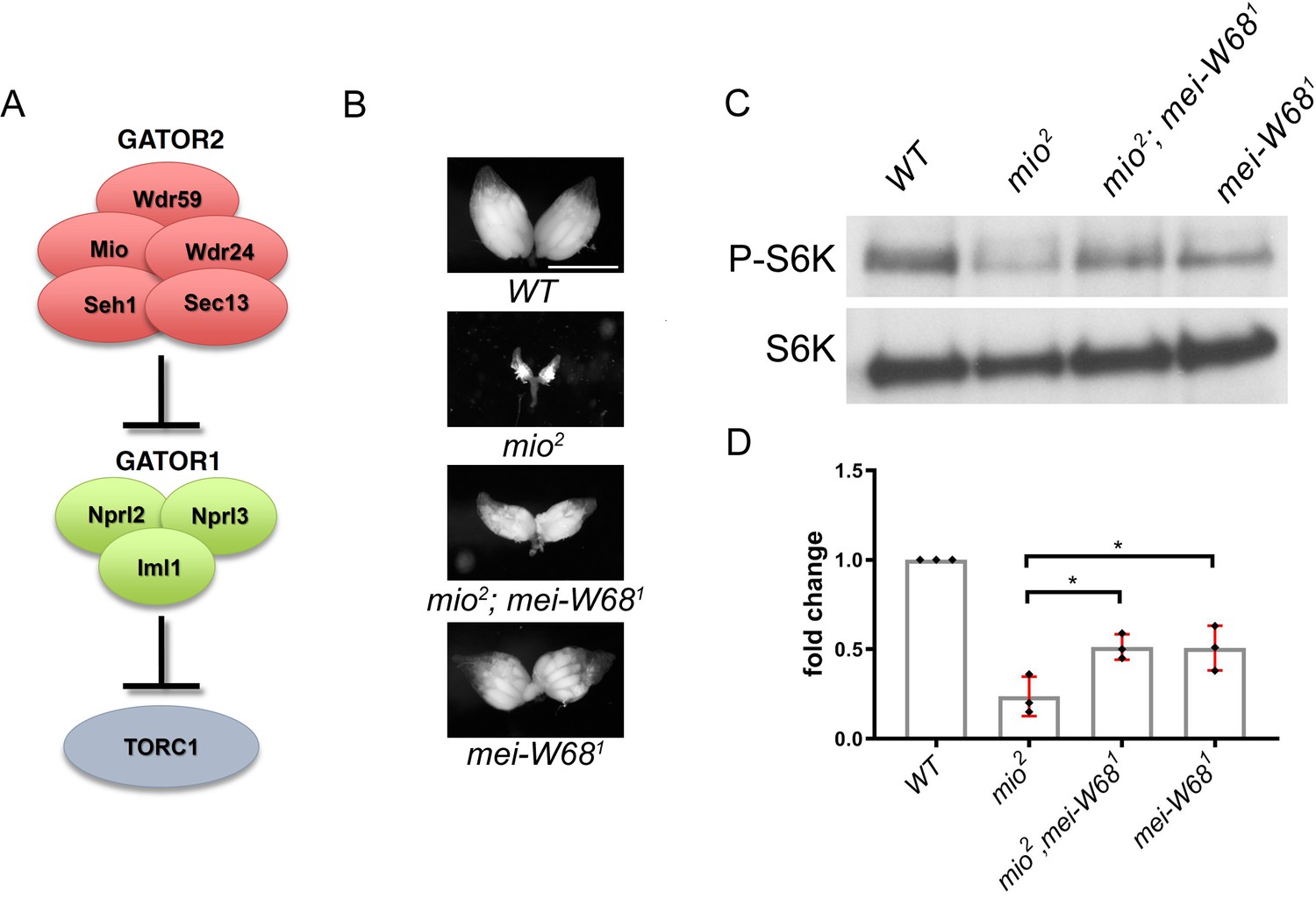{kind=link}
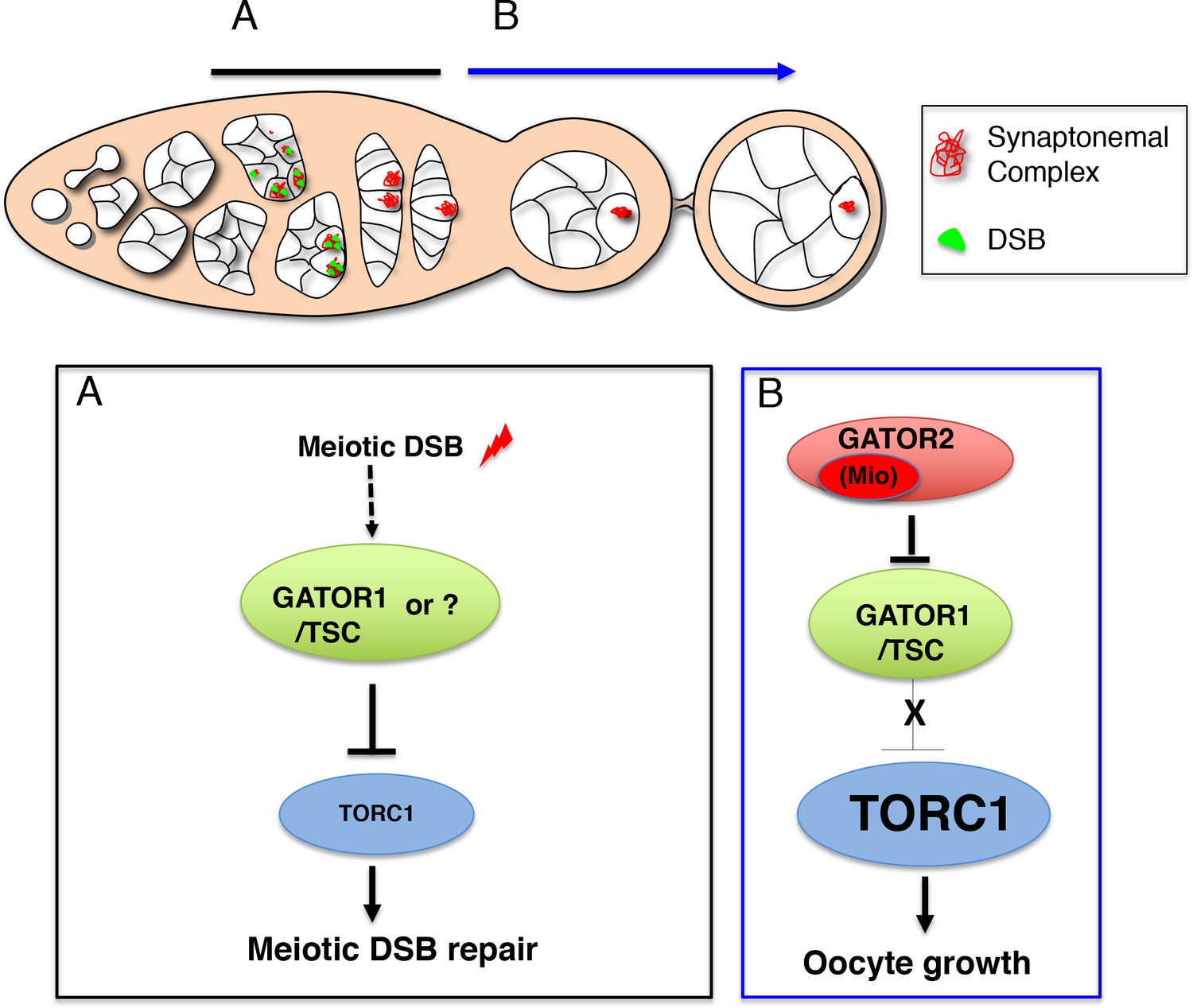{kind=link}