|
| Metamorphosis and development of the adult fly |
Formation of the adult fly
Spatial patterns of ecdysteroid receptor activation during the onset of Drosophila metamorphosis
Vesicle-mediated steroid hormone secretion in Drosophila melanogaster
Temporal profiles of nuclear receptor gene expression reveal coordinate transcriptional responses during Drosophila development
Silencing D. melanogaster lgr1 impairs transition from larval to pupal stage
The role of the prothoracic gland in determining critical weight for metamorphosis in Drosophila melanogaster
Decapentaplegic retards lipolysis during metamorphosis in Bombyx mori and Drosophila melanogaster
Dynamic feedback circuits function as a switch for shaping a maturation-inducing steroid pulse in Drosophila
Extrinsic and intrinsic mechanisms directing epithelial cell sheet replacement during Drosophila metamorphosis
Interaction between Drosophila bZIP proteins Atf3 and Jun prevents replacement of epithelial cells during metamorphosis
Juvenile hormone counteracts the bHLH-PAS transcription factors MET and GCE to prevent caspase-dependent programmed cell death in Drosophila
DPP-mediated TGFβ signaling regulates juvenile hormone biosynthesis by activating the expression of juvenile hormone acid methyltransferase
bantam miRNA promotes systemic growth by connecting insulin signaling and ecdysone production
Local requirement of the Drosophila insulin binding protein Imp-L2 in coordinating developmental progression with nutritional conditions
CDK8-Cyclin C mediates nutritional regulation of developmental transitions through the Ecdysone receptor in Drosophila
Mechanical control of whole body shape by a single cuticular protein Obstructor-E in Drosophila melanogaster
Makorin 1 regulates developmental timing in Drosophila
Coordination of cytoskeletal dynamics and cell behaviour during Drosophila abdominal morphogenesis
Snail synchronizes endocycling in a TOR-dependent manner to coordinate entry and escape from endoreplication pausing during the Drosophila critical weight checkpoint
Feedforward Regulation of Glucose Metabolism by Steroid Hormones Drives a Developmental Transition in Drosophila
The circadian clock gates Drosophila adult emergence by controlling the timecourse of metamorphosis
Drosophila E93 promotes adult development and suppresses larval responses to ecdysone during metamorphosis
Chinmo is the larval member of the molecular trinity that directs Drosophila metamorphosis
A Drosophila toolkit for HA-tagged proteins unveils a block in autophagy flux in the last instar larval fat body
Spatiotemporal organization of enteroendocrine peptide expression in Drosophila
Co-option of immune effectors by the hormonal signalling system triggering metamorphosis in Drosophila melanogaster
Neurotransmitters Affect Larval Development by Regulating the Activity of Prothoracicotropic Hormone-Releasing Neurons in Drosophila melanogaster
Proximate larval epidermal cell layer generates forces for Pupal thorax closure in Drosophila
Evaluating old truths: Final adult size in holometabolous insects is set by the end of larval development
Polycomb group genes are required for neuronal pruning in Drosophila
Effects of valproate on seizure-like activity in Drosophila melanogaster with a knockdown of Ube3a in different neuronal populations as a model of Angelman Syndrome
Single-cell transcriptomics identifies new blood cell populations in Drosophila released at the onset of metamorphosis
The macrophage genetic cassette inr/dtor/pvf2 is a nutritional status checkpoint for developmental timing
Eclosion: The pupal-adult transition
Rescheduling behavioral subunits of a fixed action pattern by genetic manipulation of peptidergic signaling
Broad specifies pupal development and mediates the 'status quo' action of juvenile hormone on the pupal-adult transformation in Drosophila
Neuromodulatory connectivity defines the structure of a behavioral neural network
Metamorphosis of memory circuits in Drosophila reveals a strategy for evolving a larval brain
Eclosion gates progression of the adult ecdysis sequence of Drosophila
Neuronal remodeling during metamorphosis is regulated by the alan shepard (shep) gene in Drosophila melanogaster
Smads and insect hemimetabolan metamorphosis
Anatomical characterization of PDF-tri neurons and peptidergic neurons associated with eclosion behavior in Drosophila
Drosophila Choline transporter non-canonically regulates pupal eclosion and NMJ integrity through a neuronal subset of mushroom body
Environmental cycles regulate development time via circadian clock mediated gating of adult emergence
Drosophila Regnase-1 RNase is required for mRNA and miRNA profile remodelling during larva-to-adult metamorphosis
Accuracy of fruit-fly eclosion rhythms evolves by strengthening circadian gating rather than developmental fine-tuning
Drosophila HNF4 directs a switch in lipid metabolism that supports the transition to adulthood
Juvenile hormone and 20-hydroxyecdysone coordinately control the developmental timing of matrix metalloproteinase-induced fat body cell dissociation
Rethinking the ecdysteroid source during Drosophila pupal-adult development
With the exception of the central nervous system, most of the structures of the adult fly develop during the larval period and take their final form during eclosion, the act of emerging from the pupal case.
The familiar external structures of the adult fly develop from imaginal discs, groups of cells that invaginate from the embryonic ectoderm, and from histoblasts and imaginal rings, cells that are set aside during embryonic development and retain their ability to divide through the larval stages. The clypeus and the labrum, two anterior head structures develop from the clypeo-labral disc. The main part of the head, including the frons, the antenna, the eyes and the maxillary palps develop from the eye-antennal disc. The proboscis develops from the labial disc. The major part of the thorax, including the wing, notum, scutellum, and pleura are formed from the wing imaginal disc, the prothorax, supporting the first leg is derived from the prothoracic disc, the legs are formed from the leg discs and the haltere is formed from the haltere disc. The abdomen is formed from nests of histoblasts that are formed as part of the embryonic epidermis.
Of the internal organs, the gut, salivary glands and trachea develop from imaginal rings that are formed during embryonic development. The larval Malpighian tubules persist into adulthood. The adult musculature develops from adepithelial cells that are attached to the imaginal discs. The gonads, ovaries and testes, develop from three sources: in females only, a nest of mesodermal follicle precursor cells, and in both males and females, genital discs and pole cells (the germ line stem cells). In the female, genital discs give rise to oviducts and uterus, and the accessory structures consisting of seminal recepticle, spermathecae, and accessory glands. In the male, the genital discs give rise to seminal vesicles, ejaculatory duct and sperm pump, penis apparatus and an accessory structure, the paragonia (Hartenstein, 1993).
The metamorphosis of the fly begins at the end of the third instar larval stage, approximately 120 hours after the beginning of embryonic development. Studies with other insects indicates that release of Ecdysone from the ring gland is triggered by the prothoracicotropic hormone, produced by four dorsolateral
neurosecretory cells of brain. Genes regulating the molting hierarchy are listed in their own site. Larval salivary gland chromosomes undergo endoreduplication and become polyploid. For information about this process, see Polytene chromosomes, endoreduplication and puffing.
Metamorphosis in Drosophila may be divided into two stages: a 12 hour prepupal period marked by pupariation (the onset of the larval-pupal transition), and a subsequent pupal period lasting 84 hours.
Pupariation is marked by a sudden release of ecdysteroid hormone secreted from the ring gland. The larval cuticle becomes the puparium or pupal case that surrounds the metamorphosing fly until it ecloses. Apolysis is the term for the retraction of the epidermis from the cuticle of the third instar larva. Once apolysis is complete, a characteristic gas bubble forms in the prepupa abdomen. At this stage the developing pupa is able to float in water.
Next the eversion of the head takes place, approximately 12 hours from the start of pupariation. The process itself is sudden, lasting about 10 minutes and orchestrated by contractions of abdominal muscles. Head eversion marks the beginning of the true pupal state. During pupariation the imaginal discs undergo eversion to form the basic shape of the adult head, thorax and abdomen. Wing, leg and haltere discs fuse to form the thorax. The eye antennal complex fuses to form a single head capsule and the head and thorax fuse with the abdomen.
During the early pupal period, final cell divisions in the wings and legs take place. Wing and leg discs inflate by the same process that drives head eversion. The next several days are marked by all the cell and tissue changes that have to take place in the development of adult structures. For example, bristles and sockets develop from precursor cells, directed by proneural genes and regulated by neurogenic genes. The layered adult cuticle develops in cycles of cuticle deposition; eye pigmentation develops and neural maturation takes place.
Early in this period a pupal to adult ecdysis takes place; for most of the pupal period the animal in the puparium is technically a pharate (cloaked or covered) adult. At the end of the pupal period eclosion (hatching) takes place, driven by an eclosion hormone (Fristrom, 1993 and Ashburner, 1989).
Ecdysteroid signaling in insects is transduced by a heterodimer of the EcR and USP nuclear receptors. In order to monitor the temporal and spatial patterns of ecdysteroid signaling in vivo, transgenic animals were established that express a fusion of the GAL4 DNA binding domain and the ligand binding domain (LBD) of EcR or USP, combined with a GAL4-dependent lacZ reporter gene. The patterns of ß-galactosidase expression in these animals indicate where and when the GAL4-LBD fusion protein has been activated by its ligand in vivo. The patterns of GAL4-EcR and GAL4-USP activation at the onset of metamorphosis reflect what would be predicted for ecdysteroid activation of the EcR/USP heterodimer. No activation is seen in mid-third instar larvae when the ecdysteroid titer is low, and strong widespread activation is observed at the end of the instar when the ecdysteroid titer is high. In addition, both GAL4-EcR and GAL4-USP are activated in larval organs cultured with 20-hydroxyecdysone (20E), consistent with EcR/USP acting as a 20E receptor. GAL4-USP activation depends on EcR, suggesting
that USP requires its heterodimer partner to function as an activator in vivo. Interestingly, no GAL4-LBD activation is observed in the imaginal discs
and ring glands of late third instar larvae. Addition of 20E to cultured mid-third instar imaginal discs results in GAL4-USP activation, but this
response is not seen in imaginal discs cultured from late third instar larvae, suggesting that EcR/USP loses its ability to function as an efficient
activator in this tissue. It is concluded that EcR/USP activation by the systemic ecdysteroid signal may be spatially restricted in vivo. GAL4-EcR functions as a potent and specific dominant negative at the onset of metamorphosis, providing a new tool for characterizing
ecdysteroid signaling pathways during development (Kozlova, 2002).
Spatially restricted and largely distinct patterns of GAL4-EcR and GAL4-USP activation were observed in the CNS at the onset of metamorphosis. Understanding the significance of these patterns will require more detailed studies that extend beyond the limits of this initial report. Nonetheless, there are several aspects of these activation patterns that are consistent with current understanding of the roles of EcR and USP in CNS development. (1) The cells where GAL4-EcR is most
active at this stage correlate with the location of the optic proliferation zones, consistent with the known role for ecdysteroids in neuronal proliferation during metamorphosis. (2) It is also interesting to note that the pattern of GAL4-USP activation in the CNS reflects a subset of the EcR-B1 expression pattern at the onset of metamorphosis. EcR-B1 is most abundantly expressed in the mushroom body neurons and surrounding cells of the optic lobes as well as the abdominal neuromeres of the ventral nerve cord. GAL4-USP activation is strongest in a cluster of cells at the anterior end of the optic lobes that could correspond to the mushroom body neurons, and is clearly elevated in the abdominal neuromeres. (3) In addition, GAL4-USP activation in the CNS is significantly reduced in an EcR mutant background, supporting the conclusion that it is acting as a heterodimer with endogenous EcR. Interestingly, low levels of GAL4-EcR activation can also be seen
in the cluster of anterior neurons in the optic lobes that show high levels of GAL4-USP activation. Unambiguous identification of these cells, however, will
require more detailed studies of the patterns of GAL4-EcR and GAL4-USP activation in the CNS as well as the use of cell-type specific markers (Kozlova, 2002).
The restricted activation of GAL4-EcR cannot be attributed to the distribution of endogenous USP in the CNS, which is widely expressed in this tissue at the onset of metamorphosis. Similarly, many neurons that express EcR-B1 in the optic lobes do not show high levels of GAL4-USP activation. One possible explanation for these limited patterns of activation is that EcR might function independently of USP in certain cells of the CNS. Alternatively, any of several possible mechanisms for the reduced levels of transactivation seen in late larval imaginal discs could account for these complex cell-type specific patterns of GAL4-LBD activation in the CNS (Kozlova, 2002).
Steroid hormones are a large family of cholesterol derivatives regulating development and physiology in both the animal and plant kingdoms, but little is known concerning mechanisms of their secretion from steroidogenic tissues. This study presents evidence that in Drosophila, endocrine release of the steroid hormone ecdysone is mediated through a regulated vesicular trafficking mechanism. Inhibition of calcium signaling in the steroidogenic prothoracic gland (PG) results in the accumulation of unreleased ecdysone, and the knockdown of calcium-mediated vesicle exocytosis components in the gland caused developmental defects due to deficiency of ecdysone. Accumulation of synaptotagmin-labeled vesicles in the gland is observed when calcium signaling is disrupted, and these vesicles contain an ABC transporter that functions as an ecdysone pump to fill vesicles. It is proposed that trafficking of steroid hormones out of endocrine cells is not always through a simple diffusion mechanism as presently thought, but instead can involve a regulated vesicle-mediated release process (Yamanaka, 2015)
This study provides several lines of evidence demonstrating that the insect steroid hormone E is secreted from the PG not by simple diffusion, but rather through a calcium signaling-regulated vesicle fusion event. Three major points come from these findings: (1) Atet, an ABCG transporter, can facilitate E passage through membranes in an ATP-dependent manner, (2) GPCR-regulated calcium signaling in the PG promotes E release, and (3) the significance of steroid hormone release by vesicle exocytosis and its implication for other steroid hormone/cholesterol trafficking processes (Yamanaka, 2015)
Atet was originally cloned in Drosophila as an ABC transporter-encoding gene with unknown function. It was found to be highly expressed in embryonic trachea, leading to its name ABC transporter expressed in trachea or Atet. In an in situ hybridization experiment, however, this study found little expression of Atet in embryonic trachea, but instead saw specific high level expression in the PG, consistent with its expression pattern in the third instar larva. Since Atet has an atypical membrane topology and can transport E across membranes in vitro, renaming this gene Atypical topology ecdysone transporter is proposed, thereby retaining the Atet gene designation (Yamanaka, 2015)
Atet belongs to the ABCG subfamily of ABC transporters, members of which in mammals have been shown to transport cholesterol as well as other steroids, such as estrogens and their metabolites, in many biological systems. The atypical membrane topology, with the N-terminal ABC domain on the non-cytoplasmic side of the membrane, has not been reported for any ABC transporter to date. However, this topology may have a strong advantage in facilitating tight control on E release by preventing Atet from functioning on the plasma membrane, due to the lack of ATP in extracellular space. This configuration therefore prevents E transport directly through the plasma membrane and confines it to a vesicle-mediated fusion process, although it requires a separate molecular mechanism to transport ATP into the secretory vesicles. This mechanism remains unclear at this point, but it may involve a specific transporter like the recently described VNUT/SLC17A9. In this context, it is interesting to note that the human Atet orthologs ABCG1 and ABCG4 are also strongly predicted by membrane topology algorithms to position their N-terminal ABC domain on the non-cytoplasmic side. These transporters mediate cellular cholesterol efflux and have recently been shown to work not on the plasma membrane but in intracellular endosomes. Clearly, additional studies on the membrane topology of ABCG transporters are warranted (Yamanaka, 2015)
The results of the RNAi screening demonstrate that CG30054, a Gαq subunit, and Plc21C, a PLCβ class enzyme, are both required for proper PG function. These findings strongly implicate the existence of an unknown GPCR and cognate ligand as mediators of the calcium signaling event that is suggested to stimulates E release from the PG. On the other hand, it is known that the PTTH receptor is Torso, a receptor tyrosine kinase and its primary role is to promote E production by inducing E biosynthetic enzyme gene transcription. These observations suggest that, at least in Drosophila, E production and release are likely regulated separately. This machinery might help the GPCR ligand to generate large pulses of steroid in a timely fashion. The identification of the GPCR as well as its ligand is necessary to further pursue this possibility (Yamanaka, 2015)
The mechanism of steroid hormone transit through lipid membranes has not been well studied and in many physiology textbooks the issue is not even discussed. When this topic is mentioned, the explanation most often given is that they can freely diffuse through lipid membranes. Despite this prevailing assumption, there are only a few reports where such transbilayer transfer of steroids by free diffusion has been analyzed. In one theoretical study, it was shown in silico that a free energy of solvation-based mechanism can produce rapid flux of estradiol, testosterone, and progesterone through a simple membrane in concordance with measured rates. However, it is well known that steroid hormone transport across membranes can indeed be an active process in some situations: there are a number of reports on transporter involvement in either uptake or elimination of steroid hormones in eukaryotes ranging from yeast to human. These reports are suggestive enough to rationalize a potential mechanism that incorporates steroid hormones into secretory vesicles, which enables regulated secretion of steroid hormones from steroidogenic tissues (Yamanaka, 2015)
Historically, the possibility of vesicle-mediated steroid hormone release has been examined using ultrastructural and biochemical approaches in multiple biological systems, including the corpus luteum in sheep. The proposed vesicle-mediated progesterone release from the sheep corpus luteum, however, was later challenged, since the peptide oxytocin was shown to be present in dense granules by immuno-EM methods and release of oxytocin and progesterone responded differently to various secretagogues. Since that time, studies investigating the possibility of vesicle-mediated steroid release in any biological system have rarely been reported. One relevant and intriguing set of studies, however, involved ultrastructural localization of E in the PG of the waxworm Galleria mellonella using immuno-EM methods. These studies suggested that E in the PG is concentrated into what appear to be secretory granules that fuse with the plasma membrane, but once again no follow up studies have been reported in the literature (Yamanaka, 2015)
In considering the various models for steroid passage through membranes, it is important to note that steroids such as progesterone, testosterone, and estradiol are significantly more hydrophobic than E. Therefore, the free energy of solvation into a lipid bilayer of E is likely to be much more positive than for sex steroids; this may preclude the use of a simple diffusion mechanism for E. In this respect, E is more similar to bile acids, which are also highly hydrophilic and need active transporters to traverse lipid bilayers. Thus, depending on their specific physiochemical properties, different steroids might use either simple passive diffusion through the plasma membrane, active transporters or some combination of these mechanisms (Yamanaka, 2015)
In summary, this work provides strong evidence that E is released from the PG by calcium-stimulated, vesicle-mediated exocytosis. Therefore, it is suggested that the prevailing 'free diffusion' model of steroid hormone secretion needs to be reconsidered. It also follows that if E uses an active export process, then the import of many hormones, in particular 20E, is also likely controlled by transporters. Given the diversity of physiological processes regulated by steroid hormones, additional characterization of the mechanisms responsible for their import and export from various cell types and tissues will have significant impact on both basic and clinical aspects of steroid hormone physiology (Yamanaka, 2015)
Many of the 21 members of the nuclear receptor superfamily in Drosophila are transcriptionally regulated by the steroid hormone ecdysone and play a role during the onset of metamorphosis, including the EcR/USP ecdysone receptor heterodimer. The temporal patterns of expression for all detectable nuclear receptor transcripts were examined throughout major ecdysone-regulated developmental transitions in the life cycle: embryogenesis, a larval molt, puparium formation, and the prepupal-pupal transition. An unexpected close temporal relationship was found between DHR3, E75B, and betaFTZ-F1 expression after each major ecdysone pulse examined, reflecting the known cross-regulatory interactions of these genes in prepupae and suggesting that they act together at other stages in the life cycle. In addition, E75A, E78B, and DHR4 are expressed in a reproducible manner with DHR3, E75B, and betaFTZ-F1, suggesting that they intersect with this regulatory cascade. Finally, known ecdysone-inducible primary-response transcripts are coordinately induced at times when the ecdysteroid titer is low, implying the existence of novel, as yet uncharacterized, temporal signals in Drosophila (Sullivan, 2003).
Total RNA was isolated from two independent collections of embryos staged at 2-h intervals throughout the 24 h of Drosophila embryonic development. Five Northern blots were prepared using equal amounts of RNA from each time point. These blots were sequentially hybridized, stripped, and rehybridized with radioactive probes derived from each of the 21 nuclear receptor genes encoded by the Drosophila genome. This approach allowed the generation of time courses of nuclear receptor gene expression that could be directly compared between family members. The transcripts detected are consistent with reported sizes. Transcripts from eight nuclear receptor genes were not detectable during embryonic development: E75C, E78, CG16801, DHR38, DHR83, dsf, eg, and svp (Sullivan, 2003).
Transcripts from nine nuclear receptor genes can be detected at the earliest time point (0-2 h): usp, EcR-A, FTZ-F1, DHR39, DHR78, DHR96, dERR, dHNF-4, and tll. This expression is consistent with the known maternal contribution of usp, EcR, and FTZ-F1. The observation that transcripts from DHR39, DHR78, DHR96, dERR, and dHNF-4 are undetectable by the next time point examined (2-4 h) suggests that these mRNAs are maternally loaded and rapidly degraded. EcR-B and usp transcripts are induced in early embryos, up-regulated at 6-8 h after egg laying (AEL), and maintain expression through the end of embryogenesis, with down-regulation of EcR-B in late embryos. EcR-A, in contrast, is expressed for a relatively brief temporal window, at 8-14 h AEL (Sullivan, 2003).
Six nuclear receptor genes are expressed in brief intervals during midembryonic stages. DHR39 and E75A are initially induced at 4-6 and 6-8 h AEL, respectively, and peak at 8-12 h AEL. This is followed by induction of DHR3, DHR4, and E75B at 8-12 h AEL, followed by ßFTZ-F1 expression at 12-18 h AEL. DHR39 appears to exhibit an expression pattern reciprocal to that of ßFTZ-F1, with lowest levels of mRNA at 14-16 h AEL and reinduction at 16-18 h as ßFTZ-F1 is repressed. This is followed by a second peak of E75A transcription at 18-22 h AEL (Sullivan, 2003).
A second group of nuclear receptors, DHR78, DHR96, dHNF-4, and dERR, is more broadly expressed at low levels throughout embryogenesis. DHR78 accumulates above its constant low level of expression between 8 and 14 h AEL. dERR exhibits an apparent mRNA isoform switch between 14 and 18 h AEL. dHNF-4 regulation also appears complex, with two size classes of mRNA induced at approximately 8-10 h AEL. While the 4.6-kb dHNF-4 mRNA is expressed throughout embryogenesis, the 3.3-kb mRNA is down-regulated at 14-16 h AEL. This timing is consistent with observations that dHNF-4 is expressed primarily in the embryonic midgut, fat body, and Malpighian tubules. Finally, nuclear receptors known to exert essential functions in patterning the early embryo, tll, kni, and knrl, are expressed predominantly during early stages (Sullivan, 2003).
Two genes that are not members of the nuclear receptor superfamily, BR-C and E74, were also examined in this study, as transcriptional markers for ecdysone pulses during development. Unexpectedly, both of these genes are induced late in embryogenesis, several hours after the rise in ecdysone titer at 6 h AEL. An approximately 7-kb BR-C transcript is induced at 10-12 h AEL and is present through the end of embryogenesis while E74B is induced at 14-16 h AEL and repressed as E74A is expressed from 16-20 h AEL. This BR-C expression pattern is consistent with the identification of the BR-C Z3 isoform in specific neurons of the embryonic CNS (Sullivan, 2003).
First-instar larvae were synchronized as they molted to the second instar, aged and harvested at 4-h intervals throughout second-instar larval development. Two Northern blots were prepared using equal amounts of total RNA isolated from a single collection of animals. Each blot was sequentially hybridized, stripped, and rehybridized to detect nuclear receptor transcription. The following transcripts were not detectable during the second instar: E75C, dERR, CG16801, DHR83, dsf, eg, svp, tll, kn, and knrl (Sullivan, 2003).
EcR-B expression is induced in mid-second-instar larvae, but does not reach maximum levels until 68-72 h AEL, just before the molt. In contrast, usp is expressed throughout the instar. A sequential pattern of nuclear receptor expression is observed that resembles the pattern seen in midembryogenesis. DHR39 and E75A are expressed in the early second instar. This is followed by induction of E75B, E78B, DHR3, and DHR4, followed by expression of ßFTZ-F1 at the end of the instar. DHR39 again shows a pattern that is approximately reciprocal with ßFTZ-F1, with highest levels during the first half of the instar. Similarly, DHR78, DHR96, and dHNF-4 exhibit broad expression patterns throughout second-instar larval development. E74A, E75A, and DHR38 are coordinately up-regulated with EcR-B at the end of the instar, between 64-72 h AEL. Finally, an approximately 9-kb BR-C transcript is detected throughout the second-larval instar (Sullivan, 2003).
Nuclear receptor gene expression was also examined throughout the third larval instar and into the early stages of metamorphosis, encompassing the ecdysone-triggered larval-to-prepupal and prepupal-to-pupal transitions. Third-instar larvae were staged relative to the molt from the second instar and harvested at 4-h intervals throughout the 48 h of the instar. Prepupae were synchronized relative to puparium formation (±15 min) and harvested at 2-h intervals up to 16 h after puparium formation (APF). Total RNA was isolated from whole animals and analyzed by Northern blot hybridization. Five blots were prepared from two independent collections of animals. These blots were sequentially hybridized, stripped, and rehybridized to detect nuclear receptor gene expression. The following transcripts were not detectable during third-instar larval or prepupal stages: CG16801, DHR83, dsf, eg, svp, tll, kn, and knrl (Sullivan, 2003).
Most nuclear receptor genes show little or no detectable expression in early and mid-third-instar larvae, a time when the ecdysone titer is low. Similar to the pattern seen in second-instar larvae, usp is expressed at relatively low levels throughout the instar and up-regulated at puparium formation, while EcR-B is induced at approximately 100 h AEL and rapidly down-regulated at puparium formation. This is followed by a sequential pattern of nuclear receptor expression similar to that seen at earlier stages. DHR39, E75A, and E78B are induced at 116-120 h AEL, in concert with the late larval ecdysone pulse, followed by maximum accumulation of E75B, DHR3, and DHR4 at 0-4 h APF. ßFTZ-F1 is expressed from 6-10 h APF, with a pattern that is approximately reciprocal to that of DHR39. EcR-A is expressed in parallel with E75B, DHR3, and DHR4 in midprepupae, similar to their coordinate expression during embryogenesis (Sullivan, 2003).
DHR78, DHR96, and dHNF-4 continue to exhibit broad expression profiles throughout third-instar larval and prepupal development. An E75 isoform not detected in embryos or second-instar larvae, E75C, is also detectable at low levels throughout most of the third instar and up-regulated in correlation with the late-larval and prepupal pulses of ecdysone. DHR38 is detectable at very low levels in early third-instar larvae, in synchrony with the early induction of E74B and BR-C. E74B is repressed, E74A is induced, and BR-C transcripts are up-regulated in late third-instar larvae, in synchrony with the late-larval ecdysone pulse. The prepupal pulse of ecdysone occurs at 10-12 h APF, marking the prepupal-to-pupal transition. EcR-A, E75A, E78B, DHR4, dERR, E75C, dHNF-4, and E74A are all induced at 10-12 h APF, in apparent response to this hormone pulse. These results are consistent with a microarray analysis of gene expression at the onset of metamorphosis where the temporal profiles of about half of these genes have been reported (Sullivan, 2003).
Most nuclear receptors can be divided into one of four classes based on this study: (1) those that are expressed exclusively during early embryogenesis (kni, knrl, tll); (2) those that are expressed throughout development (usp, DHR78, DHR96, dHNF-4); (3) those that are expressed in a reproducible temporal cascade at each stage tested (E75A, E75B, DHR3, DHR4, FTZ-F1, DHR39), and (4) those that are undetectable in these assays (CG16801, DHR83, dsf, eg, svp) (Sullivan, 2003).
Three nuclear receptor genes appear to be expressed exclusively during early embryogenesis: kni, knrl, and tll. This restricted pattern of expression fits well with the functional characterization of these genes, which have been shown to act as key determinants of embryonic body pattern. Eight genes (usp, EcR, FTZ-F1, DHR39, DHR78, DHR96, dERR, and dHNF-4) were identified that appear to have maternally deposited transcripts and thus possible embryonic functions. Indeed, maternal functions have been defined for usp, EcR, and alphaFTZ-F1 (Sullivan, 2003).
Four nuclear receptor genes are broadly expressed through all stages examined: usp, DHR78, DHR96, and dHNF-4. dHNF-4 mRNA is first detectable at 6-10 h AEL, as the ecdysone titer begins to rise. In addition, peaks of dHNF-4 expression are seen at 0, 12, and 16 h APF, in synchrony with the E74 and E75C early ecdysone-inducible genes. These observations raise the interesting possibility that this orphan nuclear receptor is regulated by ecdysone (Sullivan, 2003).
DHR38 transcripts are difficult to detect in these assays. This is consistent with studies which used RT-PCR or riboprobes for this purpose. Nonetheless, DHR38 mRNA can be detected during third-instar larval development, consistent with the widespread expression reported in earlier studies. DHR38 expression peaks at late pupal stages, consistent with its essential role in adult cuticle formation (Sullivan, 2003).
dERR and E75C display related temporal profiles of expression that do not fit with other nuclear receptor genes described in this study. Both of these genes are specifically transcribed during prepupal development, with increases in expression at 0 and 10-12 h APF. dERR, but not E75C, is also expressed during embryogenesis, with an initial induction at approximately 6 h AEL. These increases occur in synchrony with ecdysone pulses, suggesting that these orphan nuclear receptor genes are hormone inducible, although in a stage-specific manner. Further studies of dERR regulation, as well as a genetic analysis of this locus, are currently in progress (Sullivan, 2003).
Interactions between the DHR3 and E75B orphan nuclear receptors contribute to appropriate ßFTZ-F1 regulation during the onset of metamorphosis. DHR3 is both necessary and sufficient to induce ßFTZ-F1 and appears to exert this effect directly, through two response elements in the ßFTZ-F1 promoter. E75B can heterodimerize with DHR3 and is sufficient to block the ability of DHR3 to induce ßFTZ-F1. These three factors thus define a cross-regulatory network that contributes to the timing of ßFTZ-F1 expression in midprepupae. ßFTZ-F1, in turn, acts as a competence factor that directs the appropriate genetic and biological responses to the prepupal pulse of ecdysone. The patterns of DHR3, E75B, and ßFTZ-F1 expression observed at the onset of metamorphosis are consistent with these regulatory interactions as well as the expression patterns reported in earlier studies (Sullivan, 2003).
Unexpectedly, the tight linkage of DHR3, E75B, and ßFTZ-F1 expression seen at the onset of metamorphosis is recapitulated at earlier stages, after each of the major ecdysone pulses examined, in midembryogenesis and second-instar larval development. This observation suggests that the regulatory interactions between these receptors is not restricted to metamorphosis, but rather may recur in response to each ecdysone pulse during development. It is possible that this regulatory cascade contributes to cuticle deposition, which is dependent on ecdysone signaling in embryos, larvae, and prepupae. In support of this proposal, DHR3 and ßFTZ-F1 mutants exhibit defects in larval molting, suggesting that they act together to regulate this early ecdysone response (Sullivan, 2003).
Three other orphan nuclear receptor genes, E75A, DHR4, and DHR39, are expressed in concert with DHR3, E75B, and ßFTZ-F1, after the embryonic, second-instar, and third-instar ecdysone pulses. A peak of E75A expression marks the start of each genetic cascade, correlating with the rising ecdysone titer in 6- to 8-h embryos, the first half of the second instar, and in late third-instar larvae. This is followed by DHR3, E75B, and DHR4 expression which, in turn, is followed by a burst of ßFTZ-F1 expression. E78B is expressed in synchrony with DHR4 in late second and third-instar larvae, but not in embryos. These patterns of expression raise the interesting possibility that E75A, DHR4, and E78B may intersect with the cross-regulatory network defined for DHR3, E75B, and ßFTZ-F1. E75B and E78B are related to the Rev-erb vertebrate orphan nuclear receptor and are both missing their DNA binding domain. E75B and E78B null mutants are viable and fertile, suggesting that they exert redundant regulatory functions. E75A mutants die during larval stages, with no known direct regulatory targets. DHR4 mutants have not yet been described, although recent work indicates that this gene exerts essential roles in genetic and biological responses to the late larval ecdysone pulse. Further functional studies of these nuclear receptor genes should provide insight into their possible contribution to the regulatory circuit defined by DHR3, E75B, and ßFTZ-F1 (Sullivan, 2003).
Interestingly, DHR39 displays a reproducible pattern of expression that is inversely related to that of ßFTZ-F1, defining possible repressive interactions. DHR39 and ßFTZ-F1 have a similar DNA binding domain (63% identity) and bind to identical response elements, suggesting that they may exert cross-regulatory interactions. Moreover, DHR39 can repress transcription through the same response element that is activated by ßFTZ-F1. It would be interesting to determine whether the reciprocal patterns of DHR39 and ßFTZ-F1 expression during development is of functional significance (Sullivan, 2003).
The transcription of BR-C, EcR, E74, and E75 has been extensively characterized during the onset of metamorphosis, due to their rapid and direct regulation by the steroid hormone ecdysone at this stage in development. Surprisingly, however, their expression appears to be disconnected from the high-titer ecdysteroid pulses during embryonic and second-instar larval stages. As expected, EcR is induced early in embryonic development, in coincidence with the rising ecdysone titer at 4-10 h AEL, with EcR-B transcripts appearing first followed by EcR-A. BR-C mRNA, however, is not seen until 10-12 h AEL and E74B mRNA is induced even later, at 14-16 h AEL, when the ecdysteroid titer has returned to a basal level. Both EcR-B and E74B are repressed from 16-20 h AEL as E74A and E75A are induced, a switch that has been linked to the high-titer ecdysone pulse in late third-instar larvae; however, this response occurs during late embryogenesis when the ecdysteroid titer is low. A similar observation has been made for E75A expression in the Manduca dorsal abdominal epidermis, where a brief burst of E75A mRNA is detected immediately before pupal ecdysis, after the ecdysteroid titer has returned to basal levels (Sullivan, 2003).
It thus seems likely that the second instar ecdysone pulse occurs during the first half of the instar. This profile is consistent with the early induction of E75A. EcR-B and E74A, however, are not induced until the second half of the second instar, with a peak at the end of the instar. BR-C mRNA levels remain steady throughout the second instar. Finally, EcR-B, E74B, and BR-C are induced in early to mid-third-instar larvae, a time when one or more low-titer ecdysone pulses may occur. It is curious that E74B is poorly expressed relative to E74A during embryonic and second-instar larval stages, disconnecting its expression from that of EcR. This pattern is not seen in studies that focused on the onset of metamorphosis. Taken together, the temporal profiles of early gene expression (EcR, BR-C, E74, E75A) during late embryonic and late second-instar larval stages appear to be unlinked to the known ecdysteroid pulses at these stages. This could indicate that these promoters are activated in a hormone-independent manner at these stages in the life cycle. Alternatively, these ecdysone primary-response genes may be induced by a novel temporal signal that remains to be identified (Sullivan, 2003).
Several lines of evidence indicate that 20-hydroxyecdysone is not the only temporal signal in Drosophila. A major metabolite of this hormone, 3-dehydro-20-hydroxyecdysone, was shown to be as effective as 20-hydroxyecdysone in inducing target gene transcription in the hornworm, Manduca sexta. Similarly, 3-dehydro-20-hydroxyecdysone is more efficacious than 20-hydroxyecdysone in inducing Fbp-1 transcription in the Drosophila larval fat body. A high-titer pulse of alpha-ecdysone, the precursor to 20-hydroxyecdysone, can drive the extensive proliferation of neuroblasts during early pupal development in Manduca. This is the first evidence that alpha-ecdysone is responsible for a specific response in insects. It is unlikely, however, that this signal is transduced through the EcR/USP heterodimer, which shows only very low transcriptional activity in response to this ligand. Rather, recent evidence indicates that alpha-ecdysone may activate DHR38 through a novel mechanism that does not involve direct hormone binding (Sullivan, 2003).
Studies of ecdysteroid-regulated gene expression in Drosophila have also provided evidence for hormone signaling pathways that may act independently of 20-hydroxyecdysone. Several studies have identified a large-scale switch in gene expression midway through the third larval instar, an event that has been referred to as the mid-third-instar transition. It is not clear whether this response is triggered by a low-titer ecdysteroid pulse, another hormonal signal, or in a hormone-independent manner. Similarly, the let-7 and miR-125 micro-RNAs are induced at the onset of metamorphosis in Drosophila in tight temporal correlation with the E74A early mRNA, but not in apparent response to 20-hydroxyecdysone. These studies indicate that 20-hydroxyecdysone cannot act as the sole temporal regulator during the Drosophila life cycle (Sullivan, 2003).
G protein-coupled receptors (GPCRs) play key roles in a wide diversity of physiological processes and signalling pathways. The leucine-rich repeats containing GPCRs (LGRs) are a subfamily that is well-conserved through[Epub ahead of print]out most metazoan phyla and have important regulatory roles in vertebrates. This study reports on the critical role of Drosophila melanogaster LGR1, the fruit fly homologue of the vertebrate glycoprotein hormone receptors, in development as a factor involved in the regulation of pupariation. Transcript profiling revealed that lgr1 transcripts are most abundant in third instar larvae and adult flies. The tissues displaying the highest transcript levels were the hindgut, the rectum and the salivary glands. Knockdown using RNA interference (RNAi) demonstrated that white pupa formation was severely suppressed in D. melanogaster lgr1 RNAi larvae. Associated with this developmental defect was a reduced ecdysteroid titer, which is in line with significantly reduced transcript levels detected for the Halloween genes shadow (sad) and spookier (spok) in the third instar lgr1 RNAi larvae compared to the control condition (Broeck, 2014).
In Drosophila, pulsed production of the steroid hormone ecdysone
plays a pivotal role in developmental transitions such as
metamorphosis. Ecdysone production is regulated in the prothoracic gland (PG) by prothoracicotropic hormone (PTTH) and insulin-like peptides (Ilps). This study shows that monoaminergic autocrine regulation of ecdysone biosynthesis in the PG is essential for metamorphosis. PG-specific knockdown of a monoamine G protein-coupled receptor, β3-octopamine receptor (Octβ3R), resulted in arrested metamorphosis due to lack of ecdysone. Knockdown of tyramine biosynthesis genes expressed in the PG caused similar defects in ecdysone production and metamorphosis. Moreover, PTTH and Ilps signaling were impaired by Octβ3R knockdown in the PG, and activation of these signaling pathways rescued the defect in metamorphosis. Thus, monoaminergic autocrine signaling in the PG
regulated ecdysone biogenesis in a coordinated fashion on activation
by PTTH and Ilps. The study proposes that monoaminergic autocrine
signaling acts downstream of a body size checkpoint that allows
metamorphosis to occur when nutrients are sufficiently abundant (Ohhara, 2015).
In many animal species, the developmental transition is a well-known biological process in which the organism alters its body morphology and physiology to proceed from the juvenile growth stage to the adult reproductive stage. For example, in mammals, puberty causes a drastic change from adolescent to adulthood, whereas in insects, metamorphosis initiates alteration of body structures to produce sexually mature adults, a process accompanied by changes in habitat and behavior. These developmental transitions are primarily regulated by steroid hormones, production of which is regulated coordinately by developmental timing and nutritional conditions. How these processes are precisely regulated in response to developmental and environ mental cues is a longstanding question in biology (Ohhara, 2015).
In holometabolous insects, the steroid hormone ecdysone plays a pivotal role in metamorphosis. In Drosophila, metamorphic development from the third-instar larva into the adult, through the prepupa and pupa, initiates 90-96 h after hatching (hAH) at 25°C under a standard culture condition. At the onset of the larval-prepupal transition, ecdysone is produced in the prothoracic gland (PG) and then converted into its active form, 20-hydroxyecdysone (20E), in the peripheral organs. The activities of 20E terminate larval development and growth and initiates metamorphosis. Ecdysone biosynthesis is regulated in the PG by neuropeptides, enabling modulation of the timing of 20E pulses during development. The best-known stimulator of ecdysone biosynthesis is prothoracico-tropic hormone (PTTH), which is produced by neurons in the CNS. PTTH activates the receptor tyrosine kinase Torso in the PG to stimulate expression of ecdysone biosynthetic genes through the
Ras85D/Raf/MAPK kinase (MEK)/extracellular signal-regulated kinase (ERK) pathway. Insulin-like peptides (Ilps), members of another class of neuron-derived factors, activate PI3K in the PG, resulting in production of ecdysone biosynthetic proteins. The Activin/transforming growth factor-β (TGF-β) signaling pathway is also required in the PG for the expression of PTTH and Ilps receptors, although to date it remains unclear which organ produces the ligand that acts on the PG (Ohhara, 2015).
In addition to these neuropeptides, the larval-prepupal transition is modulated by environmental cues such as nutritional conditions that influence larval body size. For example, at 56 hAH, early third-instar larvae attain the minimal viable weight (MVW), at which sufficient nutrition is stored in larvae to ensure their survival through metamorphosis. After attaining MVW, larvae pass another checkpoint, critical weight (CW), defined as the minimum larval size at which starvation no longer delays the larval-prepupal transition. In Drosophila, both checkpoints occur almost simultaneously, making it difficult to distinguish them. However, CW is regarded as a body size checkpoint that initiates metamorphosis and is therefore believed to ultimately modulate ecdysone production in the PG. However, its downstream effectors and signaling pathway remain elusive (Ohhara, 2015).
Based on data obtained in Manduca and Bombyx, a G protein-coupled receptor (GPCR) has long been postulated to be essential for ecdysone biosynthesis in the PG. However, this GPCR and its ligand have not yet been identified. This study shows that monoaminergic autocrine signaling through a GPCR, β3-octopamine receptor (Octβ3R), plays an essential role in ecdysone biosynthesis to execute the larval-prepupal transition. Octβ3R is also required for activation of PTTH and Ilps signaling. It is proposed that this autocrine system acts downstream of the CW checkpoint to allow the larval-prepupal transition. It is speculated that monoamines play an evolutionarily conserved role in the regulation of steroid hormone production during developmental transitions (Ohhara, 2015).
Previously studies have shown that the GPCR Octβ3R is expressed in multiple larval tissues, including the PG. To determine whether Octβ3R is involved in ecdysone biosynthesis and metamorphosis, RNAi was used to knock down Octβ3R function specifically in the PG, using the Gal4-upstream activation sequence (UAS) system. Two different UAS-Octβ3RRNAi constructs targeting distinct regions of the Octβ3R mRNA (Octβ3RRNAi-1 and Octβ3RRNAi-2) were used to knock down Octβ3R in the PG with the help of a phantom (phm)-22-Gal4 driver. Strikingly, larvae expressing Octβ3RRNAi in the PG never developed into adult flies, and 96% of phm>Octβ3RRNAi-1 animals and 34% of phm>Octβ3RRNAi-2 animals arrested at the larval stage. When UAS-dicer2 was introduced into phm>Octβ3RRNAi-2 larvae (phm>Octβ3RRNAi-2+dicer2) to increase RNAi activity, all of these animals arrested at the larval stage. Using in situ hybridization, a significant reduction was confirmed in the Octβ3R mRNA levels in the PG of knockdown animals relative to control larvae. These data suggest that Octβ3R expression in the PG is essential for executing the larval-prepupal transition in metamorphosis (Ohhara, 2015).
Because a similar defect in the larval-prepupal transition occurs in ecdysone-deficient larvae, it was hypothesized that the Octβ3R knockdown phenotype was due to lack of ecdysone production. Consistent with this idea, the 20E titer was much lower in phm>Octβ3RRNAi-1 larvae than in control larvae just before the larval-prepupal transition (90 hAH). Moreover,
administration of 20E by feeding rescued the defect in the larval- prepupal transition caused by Octβ3R knockdown. When phm>Octβ3RRNAi-1 and phm>Octβ3RRNAi-2+dicer2 larvae were cultured on media containing 20E (1 mg/mL) from 48 hAH onward, approximately half of them developed to the prepupal stage, compared with only 2-3% of larvae not fed 20E. Thus, PG-specific loss of Octβ3R activity causes an arrest in the larval-prepupal transition due to lack of ecdysone (Ohhara, 2015).
Ecdysone is synthesized in the PG from dietary cholesterol through the action of seven ecdysone biosynthetic genes (neverland, spookier, shroud, Cyp6t3, phantom, disembodied, and shadow). Quantitative RT- PCR (qPCR) was performed to investigate whether loss of Octβ3R function affects
expression of these genes in the PG. In control larvae, expression of these genes increased dramatically between 72 and 96 hAH, when the larval-prepupal transition occurs. By contrast, in phm>Octβ3RRNAi-1 and phm>Octβ3RRNAi-2+dicer2 larvae, the expression of all of these genes was significantly reduced relative to control larvae at 96 hAH. The reduced expression of ecdysone biosynthetic genes in the PG was confirmed by in situ hybridization. Furthermore, immunostaining revealed that Neverland, Shroud, Phantom, Disembodied, and Shadow protein levels were reduced in the PG of phm>Octβ3RRNAi-1 and phm>Octβ3RRNAi-2+dicer2 larvae. Taken together, these data show that Octβ3R function is required in the PG for proper expression of ecdysone biosynthetic genes (Ohhara, 2015).
Octβ3R is thought to be activated by octopamine and tyramine binding. Octopamine is synthesized from tyramine by tyramine β-hydroxylase (Tbh), and tyramine is synthesized from tyrosine by tyrosine decarboxylase (Tdc). In Drosophila, two Tdc genes (Tdc1 and Tdc2) and one Tbh gene have been identified, and all of them are expressed in the larval CNS. Tdc1, Tdc2, and Tbh are also expressed in the PG. Furthermore, octopamine and tyramine were detected in the PG by immunostaining. Thus, octopamine and/or tyramine synthesized in the PG may activate Octβ3R in an autocrine manner to induce ecdysone production (Ohhara, 2015).
To test this, PG-specific knockdowns of Tdc1, Tdc2, and Tbh were generated. To knock down Tdc2, two constructs targeting distinct regions of the Tdc2 transcript (Tdc2RNAi-1 and Tdc2RNAi-2) were expressed along with dicer2 in the PG under the control of the phm-22-Gal4 driver (phm > Tdc2RNAi-1+dicer2 and phm > Tdc2RNAi-2+dicer2). All phm > Tdc2RNAi-1+dicer2 larvae arrested at the larval stage, and phm > Tdc2RNAi-2+dicer2 larvae were significantly delayed at the larval-prepupal transition, relative to control animals. Tdc2 mRNA level was reduced in the ring gland (RG) containing the PG in both sets of knockdown animals, as demonstrated by qPCR. Moreover, octopamine and tyramine production in the PG was impaired by Tdc2 knockdown. By contrast, Tdc1 knockdown (phm > Tdc1RNAi+dicer2) caused only a subtle delay in the larval-prepupal transition and had no detectable effect on octopamine or tyramine production. These results suggest that Tdc2 is the predominant Tdc regulating octopamine and tyramine biosynthesis in the PG and the larval-prepupal transition. Contrary to these findings, a null mutation in Tdc2 does not affect metamorphosis, and these mutant flies are viable. Thus, PG-specific knockdown causes a stronger phenotype than complete loss of Tdc2 activity in whole animals. A similar situation has been reported in regulation of metamorphosis by Activin signaling. These phenomena can be explained by a model in which some compensatory changes in other mutant tissues rescue the PG-specific knockdown phenotype in null-mutant animals (Ohhara, 2015).
PG-specific Tdc2 knockdown caused a reduction in larval 20E concentration. Therefore, whether feeding 20E to Tdc2 knockdown larvae would rescue the larval- prepupal transition defect was examined. To this end, phm > Tdc2RNAi-1+ dicer2 and phm > Tdc2RNAi-2+dicer2 larvae were cultured in media with or without 20E (1 mg/mL) from 48 hAH onward. Approximately 40% of the 20E-fed phm > Tdc2RNAi-1+dicer2 larvae developed to the prepupal stage, whereas none of those larvae grown on control media progressed beyond the larval stage. Furthermore, the delay in the larval-prepupal transition in phm > Tdc2RNAi-2+dicer2 larvae was rescued by 20E feeding. These results indicate that the defect in the larval-prepupal transition in Tdc2 knockdown animals results from a lack of 20E production. Thus, octopamine/ tyramine synthesized in the PG appears to activate Octβ3R in an autocrine manner to execute the larval-prepupal transition by regulating ecdysone production (Ohhara, 2015).
To determine which Octβ3R ligand is responsible for this autocrine signaling, Tbh was knocked down in the PG to prevent conversion of tyramine into octopamine. To knock down Tbh, two constructs targeting distinct regions of the Tbh transcript (TbhRNAi-1 and TbhRNAi-2) were expressed along with dicer2 under the control of phm-22-Gal4 (phm > TbhRNAi-1+ dicer2 and phm > TbhRNAi-2+dicer2). Although the Tbh knockdown caused a reduction in octopamine production in the PG, these larvae did not exhibit any obvious defects in the larval-prepupal transition or subsequent metamorphosi. These data suggest that tyramine, rather than octopamine, is the Octβ3R ligand that activates ecdysone production in the PG (Ohhara, 2015).
Because ecdysone biosynthesis in the PG is under the control of Ilps and PTTH signaling, it was next examined whether Octβ3R function is required to activate these signaling pathways. To detect Ilps signaling activity, a pleckstrin-homology domain fused to GFP (PH-GFP), which is recruited to the plasma membrane when insulin signaling is activated, was used. In the PG cells of control larvae, PH-GFP was only weakly localized to the plasma membrane at 48 hAH, whereas its membrane localization became increasingly evident at 60, 84, and 90 hAH. By contrast, in PG cells of phm>Octβ3RRNAi-1 larvae, the tight localization of PH-GFP to the plasma membrane was no longer detectable, indicating that activation of Ilps signaling had been disrupted. Moreover, overexpression of a constitutively active form of the Ilps receptor InR (InRCA) was able to rescue the larval arrest in phm>Octβ3RRNAi-1 animals. Next, immunostaining was performed of the diphosphorylated form of ERK (dpERK), a downstream signaling component of the PTTH pathway. dpERK expression was found to be very weak at 48 hAH, but was activated in the PG of control larvae at 60, 84, and 90 hAH; by contrast, this activation was reduced in the PG of phm>Octβ3RRNAi-1 larvae. Expression of a constitutively active form of another downstream PTTH signaling component, Ras (RasV12), rescued the larval-prepupal transition defect in phm>Octβ3RRNAi-1 animals. These results show that Octβ3R function is required to activate Ilps and PTTH signaling in the PG and that these signaling pathways execute the larval-prepupal transition. Although activation of both the Ilps and PTTH signaling pathways requires Activin/TGFβ signaling in the PG, expression of a constitutively active form of the Activin/ TGFβ receptor Baboon (BaboCA) failed to rescue the larval-prepupal transition defect in phm>Octβ3RRNAi-1 animals. This observation suggests that Octβ3R acts downstream or independent of Activin/TGFβ signaling to regulate Ilps and PTTH signaling in the PG (Ohhara, 2015).
The observations described above demonstrate that phm>Octβ3RRNAi affects Ilps and PTTH signaling in the PG as early as 60 hAH, raising the question of when Octβ3R function is required in the PG for execution of the larval-prepupal transition. To address this issue, the Gal80ts and Gal4/UAS system, which restricts expression of Octβ3R dsRNA in the PG at 18oC, but allows its expression at 28oC, was used. The results of temperature upshift and downshift experiments revealed that the larval-prepupal transition was impaired only when Octβ3R dsRNA was expressed in the PG at around 60 hAH. Notably, 60 hAH is the critical period during which larvae attain CW under nutrient-rich conditions. As noted above, when larvae are starved before attainment of CW, they are unable to transit into the prepupal stage. By contrast, starved larvae can successfully transit to prepupal/pupal stage without developmental delay once they have attained CW by growing beyond the critical period (~56 hAH) under nutrient-rich conditions in standard Drosophila medium. Thus, it is hypothesized that Octβ3R signaling acts downstream of the body-size checkpoint, or attainment of CW, to allow the larval-prepupal transition (Ohhara, 2015).
Several lines of evidence support this hypothesis. First, Octβ3R function is required for activation of Ilps and PTTH signaling detected in the PG at 60 hAH. By contrast, at 48 hAH, before the attainment of CW, neither signaling pathway is active in the PG. Second, Ilps and PTTH signaling was not activated in the PG when the larvae were starved from 48 hAH onward (early starvation), whereas these signaling pathways were active when the larvae were starved after 60 hAH (late starvation). Finally, a ligand for Octβ3R, tyramine, was detectable in the PG at 60 hAH, but decreases after this stage under a nutrient-rich condition. This decrease in tyramine was abrogated by early starvation but not by late starvation. Assuming that this decrease in tyramine in the PG is due to its secretion from PG cells, it is reasonable to propose that attainment of CW causes tyramine secretion from the PG at around 60 hAH, which in turn activates Octβ3R to regulate the Ilps and PTTH pathways, leading to the larval-prepupal transition (Ohhara, 2015).
This study demonstrates that monoaminergic regulation plays a pivotal role in ecdysone biosynthesis to induce metamorphosis and that Octβ3R acts as an upstream regulator essential for the Ilps and PTTH signaling. In addition, the data indicate that Octβ3R ligands are produced in the PG to stimulate ecdysone biosynthesis in an autocrine manner. Autocrine signaling has been proposed to mediate the community
effect, in which identical neighboring cells are coordinated in their stimulation and maintenance of cell type-specific gene expression and their differentiation, as observed in muscle development of amphibian embryos. Thus, it is proposed that monoaminergic autocrine signaling among PG cells acts to increase their responsiveness to Ilps and PTTH, thereby allowing coordinated ex- pression of ecdysone biosynthetic genes within a time window following exposure to neuropeptides (Ohhara, 2015).
These findings raise the larger question of whether monoamine acts as part of an evolutionarily conserved mechanism of steroid hormone production. In vertebrates, there is limited evidence of monoaminergic regulation of steroid hormone biosynthesis. For example, in cultured adrenal glands, catecholamine stimulates the biosynthesis of the steroid hormone cortisol in a paracrine manner to elicit a stress reaction. Another example is the Leydig cells of the mammalian testes, in which the steroid hormone testosterone is produced mainly in response to pituitary gonadotropin. However, catecholamine signaling through β-adrenergic receptors, the orthologs of Octβ3R, also promotes the production of testosterone from cultured fetal Leydig cells, which may be the site of catecholamine synthesis in the fetal and mature human testes. Thus, monoamines may play a conserved role in modulating and/or stimulating steroid hormone production during physiological and developmental transitions (Ohhara, 2015).
The timely onset of metamorphosis in holometabolous insects depends on their reaching the appropriate size known as critical weight. Once critical weight is reached, juvenile hormone (JH) titers decline, resulting in the release of prothoracicotropic hormone (PTTH) at the next photoperiod gate and thereby inducing metamorphosis. How individuals determine when they have reached critical weight is unknown. Evidence is presented that in Drosophila, a component of the ring gland, the prothoracic gland (PG), assesses growth to determine when critical weight has been achieved. The GAL4/UAS system was used to suppress or enhance growth by overexpressing PTEN or Dp110 (Pi3K92E), respectively, in various components of the ring gland. Suppression of the growth of the PG and CA, but not of the CA alone, produced larger-than-normal larvae and adults. Suppression of only PG growth resulted in nonviable larvae, but larvae with enlarged PGs produced significantly smaller larvae and adults. Rearing larvae with enlarged PGs under constant light enhanced these effects, suggesting a role for photoperiod-gated PTTH secretion. These larvae are smaller, in part as a result of their repressed growth rates, a phenotype that could be rescued through nutritional supplementation (yeast paste). Most importantly, larvae with enlarged PGs overestimated size so that they initiated metamorphosis before surpassing the minimal viable weight necessary to survive pupation. It is concluded that the PG acts as a size-assessing tissue by using insulin-dependent PG cell growth to determine when critical weight has been reached (Mirth, 2005; full text of article).
These manipulations of insulin-dependent PG growth showed that this growth is inversely related to larval growth. Suppressing the growth of the PG (P0206>PTEN - ectopically driven PTEN) produced larvae that spent more time in each instar and were larger than normal. These effects are presumably due to a combination of reduced ecdysteroid biosynthesis, which is known to delay development, and increased growth rate. Conversely, larvae with enlarged PGs (phm>Dp110; phm is a phantom GAL4 line which was used to drive expression of Dp110) showed accelerated development in the L3. Their growth rate was dependent on nutritional conditions. Whereas phm>Dp110 larvae reared on suboptimal food grew slowly, well-fed phm>Dp110 larvae grew at the same rate as controls. Together, these data indicate that the growth of the PG negatively regulates the growth rate of the whole animal and that this regulation is modulated by nutrition (Mirth, 2005).
In addition, decreasing PG size in P0206>PTEN larvae resulted in premature metamorphosis and the formation of L2 puparia. Similar L2 puparia have been described in larvae with mutations that affect the regulation of ecdysteroid biosynthesis or signaling and in larvae where the Broad isoform Z3 was overexpressed in the ring gland, resulting in its apoptosis. L2 puparia are seen in situations where ecdysone synthesis is compromised because larvae cross the threshold weight for metamorphosis prior to the production of sufficient ecdysone to initiate a larval molt, redirecting their development to the metamorphic pathway (Mirth, 2005).
Reducing PG size resulted in reduced ecdysteroid biosynthesis; P0206>PTEN larvae showed reduced ecdysteroid titers at 44 hr AEL3, and phm>PTEN larvae only molted to L2 when fed 20E. Under conditions of low ecdysteroid synthesis, fast-growing larvae could surpass the threshold for metamorphosis before the ecdysteroid titer was sufficient to induce a molt, resulting in L2 prepupae. Slower-growing larvae would be unable to reach this threshold weight before the rise in ecdysteroid titer induced the molt to L3. Indeed, undernourished, and presumably slow-growing, P0206>PTEN L2 larvae all molted to L3, whereas only 33% of the well-fed P0206>PTEN larvae molted to L3 (Mirth, 2005).
Enlarging the PG of larvae reared under constant light caused larvae to initiate metamorphosis earlier and at smaller sizes. Nevertheless, even though larvae starved early after the L3 molt were able to pupariate, they were unable to survive to pupation unless they had fed for at least 11.5 hr. This suggests that phm>Dp110 larvae starved prior to 11.5 hr AL3E initiated metamorphosis before surpassing the minimal viableweight. Furthermore, although in control larvae, critical weight and minimal viable weight are apparently attained at the same time, they are uncoupled in phm>Dp110 larvae. Therefore, the assessment of critical weight is dependent on PG growth, whereas the minimal viable weight is not (Mirth, 2005).
In Drosophila, the PGs are responsible for a size-assessment event, early in the L3, that induces the onset of metamorphosis once critical weight is surpassed. Enhancing PG growth resulted in an overestimation of body size, thereby causing the larva to initiate metamorphosis early, at a subnormal size. Under LL, the effects of enlarging the PG were enhanced, producing individuals that pupariated even earlier at even smaller sizes, suggesting that when PTTH release was unconstrained by circadian gating, the PTTH delay period was reduced. These data provide the first indication in Drosophila that the post-critical-weight PTTH release may be under photoperiod control, as it is in Manduca (Mirth, 2005).
There has been some discussion in the literature as to whether critical weight as described in Drosophila is the same as critical weight as defined in Manduca. This discussion has arisen because the definition for Manduca states that critical weight is the minimal size at which starvation can no longer delay the onset of metamorphosis, but when Drosophila larvae are starved before critical weight is reached, they die. The current data suggest that this is due to a tight relationship between minimal viable weight and critical weight in Drosophila. Effects more similar to those observed in Manduca can be obtained when pre-critical-weight Drosophila larvae are starved for an interval and then re-fed. Under these conditions, they delay metamorphosis for a period greater than the period of starvation. Much of the confusion surrounding critical weight in Drosophila has arisen because in wild-type larvae, minimal viable weight and critical weight are achieved at the same time (Mirth, 2005).
After critical weight has been surpassed, the metamorphic pathway appears to be partially suppressed by continued feeding in Drosophila. Hence, the nutrition pathway appears to promote growth and suppress metamorphosis, whereas insulin-dependent PG growth suppresses larval growth and promotes differentiation (Mirth, 2005).
The effects of increased growth in the PG are not simply due to increasing cell size, but rather are specific to the nutrition-dependent InR signaling pathway. Studies have indicated that when either dMYC or cyclinD/cdk4 are used to enlarge the PG cells, there is no reduction in overall body size. Overexpression of dMYC, of cyclinD/cdk4, and of Dp110 all enhance cell growth, but they do so in fundamentally different manners by using separate cascades. Whether the size-assessment mechanism operates via increased intracellular PIP3 levels in the PG cells or the accumulation of some other downstream component of the InR cascade in these cells is unknown (Mirth, 2005).
Although no difference in was detected ecdysteroid titers in larvae with enlarged PGs, there is evidence that increased InR signaling in the PG cells can produce mild increases in ecdysteroidogenesis and ecdysone signaling, increases that are below the level of detection of ecdysteroid-titer assays. Larvae with enlarged PGs showed both a mild increase in the transcription of phantom during feeding stages and an increase in the transcription of the early ecdysone response gene E74B. These subtle differences in ecdysteroid titers may be important for determining growth rates and for size assessment. A gradual rise in ecdysteroid titers is coincident with the time that critical weight is reached in Drosophila. Also, subtle shifts in 20E concentrations are important for growth. Basal concentrations of 20E in combination with bombyxin enhance the growth of wing imaginal tissues in vitro; slightly higher concentrations of 20E suppress growth (Mirth, 2005).
Mutations that cause imaginal disc and larval overgrowth often cause delayed pupariation and, in some cases, show low L3 ecdysteroid titers. In the case of the mutant lethal (2) giant larvae, the ring glands are smaller than normal and have the ultrastructural appearance of glands that have low rates of ecdysteroid biosynthesis. Delayed pupariation in these larvae can be rescued by implanting wild-type ring glands. Lastly, hypomorphic mutations in DHR4, a repressor of ecdysone-induced early genes, cause reductions in critical weight and early-pupariation phenotypes similar to those described in this study. Thus, the size-assessment mechanism is likely to involve surpassing a threshold ecdysteroid titer above which the activation of the ecdysone cascade occurs (Mirth, 2005).
These data allow construction of the following model for size assessment in Drosophila. As PG cells grow in response to increased InR signaling, they increase their basal level of ecdysteroid biosynthesis. Critical weight is then reached when systemic ecdysteroid concentrations surpass a threshold that sets into motion the endocrine events that will end the growth phase of larval development and allow the larva to begin metamorphosis (Mirth, 2005).
Studies in the mid-1970s defined a size-assessment event during the final instar of the moth Manduca sexta; termed critical weight, it is the minimal size required for the timely initiation of metamorphosis. How insect larvae determine when they have reached critical weight has long been a mystery. It is hypothesized that a size-assessing tissue determines when critical weight had been reached. Suppressing growth in this size-assessing tissue would cause an underestimation of body size, resulting in metamorphosis at larger than normal sizes, whereas enlarging this tissue would result in subnormal sizes. Studies in Drosophila have shown that manipulation of the growth of the PG via the InR pathway produced these types of effects. Furthermore, larvae with enlarged PGs metamorphosed at even smaller sizes when reared under LL, suggesting a role for PTTH circadian gating in this response. Smaller size arose both as a result of a reduction in growth rate, an effect that could be rescued via nutritional supplementation, and the early onset of metamorphosis. Most importantly, larvae with enlarged PGs had a remarkably reduced critical weight, suggesting that they are severely overestimating their own body size. These results offer a very new perspective on the problem of size control in insects, uniting the recent data exploring the role of nutrition and the insulin-receptor pathway on growth with the classical physiological experiments that defined critical weight (Mirth, 2005).
Insect morphogen Decapentaplegic (Dpp) functions as one of the key extracellular ligands of the Bone Morphogenetic Protein (BMP) signaling pathway. Previous studies in insects mainly focused on the roles of Dpp during embryonic development and the formation of adult wings. This study demonstrated a new role for Dpp in retarding lipolysis during metamorphosis in both Bombyx mori and Drosophila melanogaster. CRISPR/Cas9-mediated mutation of Bombyx dpp causes pupal lethality, induces an excessive and premature breakdown of lipids in the fat body, and upregulates the expressions of several lipolytic enzyme genes, including brummer (bmm), lipase 3 (lip3), and hormone-sensitive lipase (hsl), and lipid storage droplet 1 (lsd1), a lipid droplets (LD)-associated protein gene. Further investigation in Drosophila reveals that salivary gland-specific knockdown of the dpp gene and fat body-specific knockdown of Mad involved in Dpp signaling phenocopy the effects of Bombyx dpp mutation on pupal development and lipolysis. Taken together, these data indicate that the Dpp-mediated BMP signaling in the fat body maintains lipid homeostasis by retarding lipolysis, which is necessary for pupa-adult transition during insect metamorphosis (Qian, 2023).
Antagonistic actions of ecdysone and of insulins, acting through Foxo and 4E-BP, determine final size in Drosophila
All animals coordinate growth and maturation to reach their final size and shape.
In insects, insulin family molecules control growth and metabolism, whereas pulses of the steroid 20-hydroxyecdysone (20E) initiate major developmental transitions. 20E signaling also negatively controls animal growth rates by impeding general insulin signaling involving localization of the transcription factor dFOXO and transcription of the translation inhibitor 4E-BP. The larval fat body, equivalent to the vertebrate liver, is a key relay element for ecdysone-dependent growth inhibition. Hence, ecdysone counteracts the growth-promoting action of insulins, thus forming a humoral regulatory loop that determines organismal size (Colombani, 2005).
In metazoans, the insulin/IGF signaling pathway (IIS) plays a key role in regulating growth and metabolism. In Drosophila, a family of insulin-like molecules called Dilps activates a unique insulin receptor (InR) and a conserved downstream kinase cascade that includes PI3-kinase (PI3K) and Akt/PKB. Recent genetic experiments have established that this pathway integrates extrinsic signals such as nutrition with the control of tissue growth during larval stages. The larval period is critical for the control of animal growth, since it establishes the size at which maturation occurs and, consequently, the final adult size. Maturation is itself a complex process that is controlled by the steroid 20-hydroxyecdysone (20E). Peaks of 20E determine the timing of all developmental transitions, from embryo to larva, larva to pupa, and pupa to adult. Ecdysteroids are mainly produced by the prothoracic gland (PG), a part of a composite endocrine tissue called the ring gland. Final adult size thus mainly depends on two parameters: the speed of growth (or growth rate), which is primarily controlled by IIS, and the overall duration of the growth period, which is limited by the onset of the larval-pupal transition and timed by peaks of ecdysone secretion. Very little is known concerning the mechanisms that coordinate these two parameters during larval development (Colombani, 2005).
To investigate the function of ecdysone in controlling organismal growth, a genetic approach was developed that allowed modulation basal levels of ecdysteroids in Drosophila. The initial rationale was to modify the mass of the ring gland in order to change the level of ecdysteroid production. For this goal, the levels of PI3- kinase activity were manipulated in the PG by crossing P0206-Gal4 (P0206>), a line with specific Gal4 expression in the PG and corpora allata (CA), with flies carrying UAS constructs allowing expression of either wild-type (PI3K) or dominant-negative (PI3KDN) PI3-kinase. As expected, these crosses produced dramatic autonomous growth effects in the ring gland, and particularly in the PG: tissue size was increased upon PI3K activation and decreased upon inhibition. Surprisingly, the changes in ring gland growth were accompanied by opposite effects at the organismal level. P0206>PI3K animals (with large ring glands) showed reduced growth at all stages of development and produced emerging adults with reduced size and body weight (78% of wt). Conversely, reducing PI3K activity in the ring gland of P0206>PI3KDN animals led to increased growth and produced adults with 17% greater weight on average. Adult size increase was attributable to an increase in cell number in the wing and the eye. Adult size reduction was accompanied by a decrease in cell number in the wing and in cell size in the eye (Colombani, 2005).
Importantly, the timing of embryonic and larval development of these animals was comparable to control. Both the L2 to L3 transition as well as the cessation of feeding (wandering) occurred at identical times. Further, animals entered pupal development at the same time, except for P0206>PI3KDN animals, which showed a 1-2 hrs delay intrinsic to the UAS-PI3KDN line itself. The duration of pupal development was slightly modified, however, as adult emergence was delayed in P0206>PI3K animals and advanced in P0206>PI3KDN animals, albeit by less than 4 hours following 10 days of development. In contrast, the speed of larval growth was found to be increased in P0206>PI3KDN animals and decreased in the P0206>PI3K animals background at the earliest stage that could be measured (early L2 instar). Because none of these effects were observed when PI3K levels were modified specifically in the CA using the Aug21- Gal4 driver, it was concluded that the observed phenotypes are solely due to PI3K modulation in the PG. Together, these results demonstrate that manipulating PI3K levels in the PG induces non-autonomous changes in the speed of larval growth (growth rate effects), without changing the timing of larval development (Colombani, 2005).
To investigate whether these effects could be attributed to changes in 20E levels, ecdysteroid titers were measured in third instar larvae of the different genotypes. Early after ecdysis into third instar (74hrs AED) ecdysteroids are present at basal level. They accumulate to an intermediate plateau around 90hrs AED and reach peak levels before pupariation (120hrs AED). Because early L3 levels are below the detection limit of the EIA assay, ecdysteroid titers were measured at the intermediate plateau (90hrs AED). In these conditions, a very modest increase of ecdysteroids was observed in P0206>PI3K animals larvae and a small but significant decrease in P0206>PI3KDN animals animals. This was further confirmed by measuring the transcript levels of a direct target of 20E, E74B, which responds to low/moderate levels of 20E. However, in early L3 larvae with basal ecdysteroid levels (74hrs AED), differences in E74B transcripts were clearly visible, with a 1.9-fold increase seen for P0206>PI3K animals and a 1.7-fold decrease for P0206>PI3KDN animals. This establishes that basal circulating levels of 20E are modified in response to manipulation of PI3K levels in the PG. It also suggests that the differences observed on basal 20E level off with the strong global increase of ecdysteroids in mid/late L3 (Colombani, 2005).
Several related lines of evidence strengthen these results: (1) the increase in growth rate and size observed in P0206>PI3KDN animals can be efficiently reverted by adding 20E to their food; (2) feeding wild-type larvae 20E recapitulates the effects observed in P0206>PI3K animals animals; (3) ubiquitous silencing of EcR using an inducible EcR RNAi construct results in a growth increase similar to that observed in P0206>PI3KDN larvae. Finally, the phantom (phm) and disembodied (dib) genes, which are specifically expressed in the PG and encode hydroxylases required for ecdysteroid biosynthesis, showed 1.65- and 2.2- fold increased expression, respectively, upon PI3K activation in the ring gland. This supports the notion that 20E biosynthesis is mildly induced in these experimental conditions. In line with previous results, neither 20E treatment nor EcR silencing has any effect on developmental timing. Overall, the results indicate that manipulating basal levels of 20E signaling in various ways modifies the larval growth rate without affecting the timing of the larval transitions (Colombani, 2005).
Variations in ecdysone levels in animals with different sized ring glands could be due to changes in the PG tissue mass or, alternatively, to a specific effect of PI3K signaling in the secreting tissue. To distinguish between these two possibilities, extra growth was induced in the PG using either dMyc or CyclinD/Cdk4, two potent growth inducers in Drosophila. Although the size of the larval ring gland was markedly increased under these conditions, no effect on pupal or adult size was observed, implying that the size of the ring gland is not the critical factor in the control of body size. Instead, it is likely that the InR/PI3K signaling pathway can specifically activate ecdysone production from the PG (Colombani, 2005).
The possibility was tested that ecdysone signaling opposes the growth-promoting effects of IIS in the larva. To test this, larvae were fed 20E and xPI3K activity was assessed in vivo using a GFP-PH fusion (tGPH) as a marker. Membrane tGPH localization shows a marked decrease in the fat body of 20E-fed animals, and this effect can be reverted by specifically silencing EcR in the fat body. This indicates that ecdysone-induced growth inhibition correlates with decreased IIS, and is mediated through the nuclear receptor EcR. Conversely, larvae with PI3KDN expression in the PG show a 4.2-fold increase in the global levels of dPKB/Akt activity, as measured by the phosphorylation levels of serine 505. In Drosophila cells (as in other metazoan cells) high levels of PI3K/AKT activity cause the transcription factor dFOXO to be retained in the cytoplasm, while low PI3K/AKT activity allows dFOXO to enter the nucleus where it promotes 4E-BP transcription. In larvae with ectopic PI3K expression in the PG, a strong increase is observed in nuclear dFOXO in fat body cells. Similar results were obtained by feeding larvae with 20E. Conversely, inactivation of EcR signaling in fat body cells was carried out using the clonal over-expression of a dominant-negative form of EcR (EcRF645A). In these conditions, a reduction was observed in the accumulation of dFOXO in the nuclei of EcRF645A-expressing cells. As an expected consequence of the increased nuclear dFOXO, global accumulation of 4E-BP transcripts was consistently higher in P0206>PI3K animals as well as in 20E-fed early L3 larvae as compared to control animals, and reduced in arm>EcR-RNAi animals. Together, these results indicate that ecdysone-dependent inhibition of larval growth correlates with a general alteration of insulin/IGF signaling, and a relocalization of dFOXO into the cell nuclei. To more directly test the role of dFOXO in the growth-inhibitory function of ecdysone signaling, the effects of increasing ecdysone levels were examined in a dFOXOmutant genetic background. Although homozygous dFOXO21 animals do not display a detectable growth phenotype, introducing the dFOXO21 mutation was sufficient to totally revert the growth defects of P0206>PI3K animals animals, either when homozygous or heterozygous. This data establishes that dFOXO is required for 20E-mediated growth inhibition (Colombani, 2005).
The endocrine activities of the brain and the fat body have previously been implicated in the humoral control of larval growth. In order to test for possible roles of these two organs in mediating the systemic growth effects of ecdysone, EcR expression was silenced specifically in the brain cells that produce insulins (IPCs) or in the fat body. While specific expression of EcR RNAi in the IPCs fails to reproduce the overgrowth observed in armGal4>EcR-RNAi animals, EcR silencing in the fat body elicits an acceleration of larval growth and a remarkable increase in pupal size. Moreover, no detectable delay in the larval timing was observed in pplGal4>EcR-RNAi animals. Thus, specifically reducing 20E signaling in the fat body is sufficient to recapitulate the systemic effects of global EcR silencing. This demonstrates that the fat body is a major target for ecdysone, and that this tissue can act to relay the 20E growth-inhibitory signal to all larval tissues (Colombani, 2005).
In summary, these results establish an additional role for 20E in modulating animal growth rates. This function is mediated by an antagonistic interaction with IIS that ultimately targets dFOXO function. A similar antagonistic interaction between 20E and insulin signaling controls developmentally-regulated autophagy in Drosophila larva (Colombani, 2005).
Although a direct effect of ecdysone on the cellular growth rate of all larval tissues cannot be ruled out, the experiments reveal a key role for the fat body in relaying ecdysone-dependent growth control signals. Together with previous work, these data suggest that various inputs such as nutrition and ecdysone converge on this important regulatory organ, which then controls the general IIS to modulate organismal growth (Colombani, 2005).
Histoblast nest spreading initiates with the projection of leading comet-like protrusions, followed by apical cytoskeletal activity and active crawling over the underlying basal membrane, and terminates with the implementation of an apparent purse string, reminiscent of those described during dorsal closure, C. elegans ventral enclosure or wound healing (Ninov, 2007).
The comet-like protrusions of guiding histoblasts break through the LEC epithelial barrier, leading to planar intercalation of histoblast cell bodies. They account for the capacity of histoblasts to achieve migration within the bounded epithelial layer. Indeed, electron micrographs reveal that the advancing histoblasts form junctions with non-adjacent LECs before the adjacent LECs histolyze, thus insuring the continuity of the epidermis. Time-lapse observations suggest that these protrusions grow by sequential addition of actin molecules at their forward end. In this sense, they resemble, although being considerably slower, the actin tails employed by Listeria to propel through the cytoplasm of infected cells, or the actin-rich pseudopodia extended by neutrophils in response to chemoattractants. Proper actin cytoskeleton dynamics appear to be essential to build up these protrusions and the full repertoire of activities leading to the expansion of histoblast nests. The equilibrium between actin polymerization and depolymerization activities should be exquisitely regulated, and the forced polymerization of actin by Profilin overexpression not only blocks the cytoskeletal dynamics of single cells, but impedes the spreading of the whole histoblast nest. Potential roles for further actin dynamics regulators, the Arp2-Arp3 (Arp14D-Arp66B - FlyBase) complex, Dynamin (Shibire), membrane polyphosphoinositides, Cdc42, WASp-family proteins and other molecules in building up these projections remain to be explored. Further, although these protrusions appear to have a mechanical role, they also seem to be involved in the recognition of guidance cues, as they follow stereotyped paths. Indeed, gradients of cell affinity have been described for the patterning of the Drosophila abdomen, and it would be of major interest to understand how these cells interpret the larval landscape (Ninov, 2007).
The mechanisms involved in the death of LECs have been a matter of debate. While ultrastructural analysis suggests that LECs are phagocytosed, other studies suggested that LECs are histolyzed and die by autophagy. The current findings are conclusive in this respect. The death of LECs involves a caspase-mediated apoptotic process that implicates cytoskeletal remodelling and apical cellular constriction leading to delamination. The actomyosin mediated contractile force of dying LECs contributes in bringing together neighbouring histoblasts. Once the LECs initiate extrusion, they become immediate targets for circulating haemocytes, which extend membrane projections and engulf them. Finally, LECs are degraded inside haemocytes (Ninov, 2007).
Apical constriction is a process shared by multiple morphogenetic events, e.g., Drosophila mesodermal cells accumulate myosin and apically constrict during gastrulation under the control of the small GTPase Rho. Myosin activity is also sufficient to promote the apical constriction and elimination of photoreceptor cells in the Drosophila eye in response to the overexpression of an activated form of the Rok kinase. Indeed, this study found that the apical contractility of LECs depends on the level of phosphorylation of the MRLC and could be enhanced or abolished by modulating the counteracting kinase and phosphatase activities of Rok and MLCP. As a consequence, LEC delamination is either accelerated or delayed. How these regulatory activities are themselves regulated remains to be established. Yet, the LEC extrusion defects observed in weakened caspase cascade conditions after P35 overexpression strongly suggest that apoptotic signals could be involved in the trigger of actomyosin contractility in LECs. Apical contraction would thus be an early event in the LEC apoptotic process. Being particularly important to analyse the differences that modulate the activity of myosin during apical constriction of living cells and during extrusion of apoptotic cells, the replacement of LECs could become an exceptionally suitable model to unravel how myosin activity is regulated in apoptotic cells in vivo (Ninov, 2007).
The recruitment of haemocytes to dying LECs during abdominal cell replacement is extremely fast. The apical constriction of LECs takes about 2 hours, but the time that a haemocyte needs to fully engulf a LEC is less than 10 minutes. This entails a very reliable chemoattracting mechanism. In mammals, caspase 3-dependent lipid attraction signals, released by dying cells, induce the migration of phagocytes. Furthermore, several receptors are implicated in corpse recognition, including lectins, integrins, tyrosine kinases, the phosphatidylserine receptor (PSR) and scavenger receptors. In Drosophila, the elements involved in cell recognition by macrophages are mostly unknown. Haemocytes express Croquemort, a scavenger receptor homologue, which is required for the uptake of dead cells, and Pvr, a homologue of the vertebrate PDGF/VEGF receptor that seems to affect their motility. Still, the signals that haemocytes recognize in dying cells and the links between those signals and the apoptotic cascade are essentially unknown (Ninov, 2007).
As macrophages are responsible for much of the engulfment of dead cells in developing animals, an important role for macrophages in tissue morphogenesis has been suggested. However, this is not the case during abdominal morphogenesis, as the inhibition of haemocyte motility, which abrogates the removal of LECs, does not affect their replacement by histoblasts. These results are consistent with studies showing that macrophage removal of cell debris is not required for the regeneration of laser-induced wounds in Drosophila (Ninov, 2007).
Histoblast nest expansion is tightly coordinated with LEC removal. A naive view of the process of LEC extrusion suggests that their death is altruistic - it would promote the expansion of histoblasts. However, several results suggest that LECs do not execute this process autonomously. First, histoblast nests initiate their expansion in the absence of LEC death. Second, histoblast nests, during their spreading, grow, with no obvious planar orientation, by stochastic cell divisions not restricted to their edges. Finally, and most importantly, the inhibition of histoblast proliferation exerts non-autonomous effects on both extrusion and removal of LECs. A working model in which histoblast proliferation and LEC death are synchronized by a spatially and temporally controlled exchange of signals (secreted ligands or cell-to-cell communication modules) is thus strongly appealing. This potential mechanism for replacement of LECs by histoblasts somewhat resembles the elimination and death by anoikis of amnioserosa cells upon dorsal closure completion during Drosophila embryogenesis. Through this process, physical contacts and intracellular signalling among epithelial leading cells, the amnioserosa and the yolk sac coordinate the different behaviour of these cell types, which is essential for the accurate progress of both germ band retraction and dorsal closure. In this scenario, coordinated extrinsic and intrinsic events, hormonal inputs, cell contacts and cell signalling events will be responsible for the ordered proliferation and expansion of histoblasts and the extrusion and death of LECs (Ninov, 2007).
An alternative mechanism for the ordered cell substitution taking place during abdominal morphogenesis involving cell competition could also be proposed. Competition can be defined as an interaction between individuals brought about by a shared requirement leading to a reduction in the survivorship, growth and/or reproduction rates. Classical experiments in Drosophila imaginal discs have shown that cells heterozygous mutant for ribosomal protein genes (Minutes) placed beside wild-type cells are outcompeted and eliminated from the epithelium. More recent work has shown that imaginal wild-type cells are outcompeted by cells with growth advantage overexpressing the proto-oncogene Myc. Cell competition does not just apply to the fight for survival of cells with their 'fitness' experimentally altered, but also applies to the homeostasis of self-renewing cell pools such as lymphocytes or stem cells. The substitution of LECs by histoblasts closely resembles cell competition. Rapidly dividing and expanding histoblasts may become competent to displace the surrounding less-metabolically-active LECs. During normal development, having 'weaker' neighbours, histoblasts do not compete against each other, and cells from Minute clones in the abdomen are not eliminated in heterozygous animals. However, when confronted with death-resistant LECs, 'winner' histoblasts may become 'losers'. Histoblasts in an increasingly crowded environment will compete against each other, and the less fit individuals (less competent in signalling reception and transduction, or with slower proliferation rates) would eventually become more sensitive to 'killing' signals and would die (Ninov, 2007).
These findings demonstrate that the replacement of LECs by histoblasts, independently of being driven by cooperative mechanisms, cell competition or both, represents an extremely amenable morphogenetic model for the study of the dynamic control of the cell cycle and cell death, of the coordination of cytoskeleton activities and cell adhesion, and for the study of cell invasiveness (Ninov, 2007).
Epithelial sheet spreading and fusion underlie important developmental processes. Well-characterized examples of such epithelial morphogenetic events have been provided by studies in Drosophila, and include embryonic dorsal closure, formation of the adult thorax and wound healing. All of these processes require the basic region-leucine zipper (bZIP) transcription factors Jun and Fos. Much less is known about morphogenesis of the fly abdomen, which involves replacement of larval epidermal cells (LECs) with adult histoblasts that divide, migrate and finally fuse to form the adult epidermis during metamorphosis. This study implicates Drosophila Activating transcription factor 3 (Atf3), the single ortholog of human ATF3 and JDP2 bZIP proteins, in abdominal morphogenesis. During the process of the epithelial cell replacement, transcription of the atf3 gene declines. When this downregulation is experimentally prevented, the affected LECs accumulate cell-adhesion proteins and their extrusion and replacement with histoblasts are blocked. The abnormally adhering LECs consequently obstruct the closure of the adult abdominal epithelium. This closure defect can be either mimicked and further enhanced by knockdown of the small GTPase Rho1 or, conversely, alleviated by stimulating ecdysone steroid hormone signaling. Both Rho and ecdysone pathways have been previously identified as effectors of the LEC replacement. To elicit the gain-of-function effect, Atf3 specifically requires its binding partner Jun. These data thus identify Atf3 as a new functional partner of Drosophila Jun during development (Sekyrova, 2010).
Metamorphosis of Drosophila larvae into pupae and adult flies provides remarkable examples of morphogenetic changes that involve replacement of entire cell populations. Epithelia that had served larval function undergo programmed cell death while imaginal cells proliferate and differentiate to take their position. The Drosophila abdomen is an attractive system for studying the developmental replacement of one epithelial cell population with another. Unlike the adult head and thorax with appendages, all forming from pre-patterned imaginal discs, the adult abdomen derives from histoblasts that reside in each abdominal segment. Soon after the onset of metamorphosis, the diploid histoblasts undergo an initial phase of synchronized cell divisions; later the histoblasts expand while proliferating and replace the old polyploid larval epidermal cells (LECs) that cover the surface of the abdomen. To free space for the histoblasts, LECs are extruded from the epithelial monolayer. In order to maintain integrity of the epithelia, changes in cell adhesion and cell migration must be precisely orchestrated during this tissue remodeling (Sekyrova, 2010).
Rho kinase signaling, which stimulates constriction of the apical actomyosin cytoskeleton through myosin phosphorylation, is necessary for the extrusion and the ensuing apoptosis of LECs. Perturbed myosin phosphorylation leaves the process of the epithelial exchange incomplete, with residual LECs obstructing closure of the adult abdominal epidermis at the dorsal midline. A similar defect results from compromised function of the ecdysone receptor (EcR), which is required for both the initial phase of histoblast proliferation and for the removal of LECs. Other factors besides Rho signaling and EcR that regulate the epithelial cell replacement are unknown (Sekyrova, 2010).
This study implicates Atf3 (Activating transcription factor 3 -- FlyBase), the single Drosophila ortholog of the vertebrate Activating transcription factor 3 (ATF3) and Jun dimerization protein 2 (JDP2) in abdominal development. ATF3 and JDP2 belong among basic region-leucine zipper (bZIP) proteins, some of which play important roles in epithelial morphogenesis. Particularly the functions of Jun and Fos bZIP proteins in epithelial closure events during development are well understood owing to genetic studies in Drosophila. By contrast, no morphogenetic function has yet been reported for Atf3 in Drosophila (Sekyrova, 2010).
Mammalian ATF3 and JDP2 form homodimers but preferentially dimerize with members of the Jun subfamily (Aronheim, 1997; Hai, 1989; Hsu, 1991), functioning either as transcriptional activators (ATF3-Jun) or repressors (JDP2-Jun). Based mainly on cell-culture studies, multiple roles in cell proliferation, differentiation and apoptosis have been ascribed to ATF3 and JDP2. Atf3-/- mice are viable but suffer from altered glucose and immune homeostasis. Also Jdp2-/- mice survive but produce extra fat in their brown adipose tissue. In vivo significance of the interaction between the ATF3 or JDP2 proteins and Jun remains unclear (Sekyrova, 2010).
This study shows that Atf3 interacts biochemically and genetically with Jun in Drosophila. Temporal downregulation of atf3 transcription during metamorphosis is crucial, since sustained atf3 expression alters adhesive properties of LECs, thus preventing their extrusion and replacement by the adult epidermis. This effect of Atf3 requires the presence of Jun (Sekyrova, 2010).
Among Drosophila bZIP proteins, the predicted product of the CG11405 gene (Activating transcription factor 3), located on the X chromosome, shows the closest similarity to the mammalian ATF3 and JDP2 proteins. The DNA-binding/dimerization bZIP domains of the human ATF3 and Drosophila Atf3 proteins are identical in 60% of their amino acids; there is 58% identity between Atf3 and JDP2 in this region (Sekyrova, 2010).
Dimerization between Atf3 and Jun in Drosophila has been theoretically predicted and confirmed by a yeast two-hybrid screen. To demonstrate direct binding, co-immunoprecipitation experiments were conducted. The endogenous Jun protein from Drosophila S2 cells co-precipitates with a transiently expressed Atf3 whereas Fos did not. A DNA mobility-shift assay with recombinant bZIP domains of Atf3, Jun and Fos was conducted to test for their DNA-binding properties. Atf3 specifically bound an ATF/CRE consensus element but not the AP-1 site, which was recognized by the Jun-Fos (AP-1) complex. Although Atf3 bound DNA by itself, presumably as a homodimer, the binding was enhanced in the presence of Jun. Fos did not synergize with Atf3 in DNA binding. Excess unlabeled DNA bearing the ATF/CRE binding site competed for the Atf3 bandshift activity whereas the AP-1 binding element did not. These results have shown that, like ATF3 or JDP2 in mammals, Atf3 in Drosophila selectively dimerizes with Jun, with which it cooperatively and specifically binds the ATF/CRE DNA element (Sekyrova, 2010).
To test whether Atf3 and Jun interact in vivo, experiments were conducted in the Drosophila compound eye, the precise structure of which sensitively reflects genetic interactions. Overexpression of atf3 under the GMR-Gal4 driver disrupted the ommatidial arrangement, resulting in smaller eyes with a glossy appearance. This atf3 misexpression phenotype could be completely suppressed by simultaneous RNAi-mediated knockdown of jun but not of fos. Conversely, the phenotype was exacerbated when jun was overexpressed in the eye together with atf3, suggesting that it is Atf3 in a complex with Jun that derails the normal eye development. Neither RNAi nor overexpression of jun alone had any effect on eye morphology. Interestingly, like depletion of Jun, co-expression of fos under the GMR-Gal4 driver completely averted the atf3 misexpression phenotype, restoring the normal appearance of the eye. Expression of fos or its mutant versions alone had no effect. These data can be explained by the ability of the surplus Fos to bind Jun and thus reduce its availability for interaction with the Atf3 protein. This interpretation is further supported by experiments showing that expression of the truncated bZIP domain of Fos is sufficient to suppress the Atf3 gain-of-function phenotype, whereas its transcription activation domain or phosphorylation sites are dispensable (Sekyrova, 2010).
Taken together, these results show that Atf3 cooperates with Jun, as Jun is specifically required for an effect caused by overexpression of Atf3 in the developing eye. Given the capacity of both Atf3 and Fos to bind Jun, and based on the ability of Jun to enhance and of Fos to suppress the Atf3 gain-of-function phenotype, it is suggested that Atf3 and Fos compete for their common partner Jun in vivo (Sekyrova, 2010).
To find out whether Atf3 is required for Drosophila development and whether its absence might resemble a phenotype caused by loss of its partner Jun, atf3 mutant flies were generated. The longest deletion (line atf376) obtained by imprecise excision of a P element, removed the entire bZIP domain of Atf3, and atf376 hemizygous (male) larvae lacked detectable atf3 mRNA. Thus, atf376 probably represents a null allele. Most atf376 larvae die soon after hatching and during all three larval stages. Only a few (approximately 2%) reach the third instar but die before metamorphosis as defective pseudopuparia. Expression of atf3 cDNA under the ubiquitous armadillo (arm-Gal4) driver rescued some atf376 hemizygotes to adults, confirming that loss of atf3 was the cause of the lethal phenotype. Interestingly, the moribund atf376 larvae abnormally enlarged lipid droplets in their fat body, thus displaying a phenotype reminiscent of that in mice lacking one of the Atf3 orthologs, JDP2. However, in contrast to viable Jdp2 or Atf3 knockout mice, atf3 is an essential gene in Drosophila (Sekyrova, 2010).
Fly embryos lacking the function of Jun or Fos die because of the failed dorsal closure. However, atf376 embryos develop normally, without the dorsal open defect, even when derived from atf3-deficient germline clones induced in atf376/ovoD1 mothers. Thus, unlike its partner Jun, Atf3 is not required for dorsal closure, suggesting that dorsal closure is regulated by Jun-Fos dimers and that the Atf3-Jun complex has another function later in development (Sekyrova, 2010).
Consistent with the vital requirement for Atf3 during larval stages, atf3 mRNA was expressed in embryos and larvae. Expression then sharply declines by the late-third larval instar, and no atf3 mRNA was detected by northern blot hybridization in wandering larvae and during metamorphosis from the time of puparium formation until the second day of pupal development. Detailed RT-PCR analysis showed that atf3 downregulation coincided with the cessation of feeding and the onset of metamorphosis [0 hours after puparium formation (APF)]. A pulse of expression occurred at 6 hours APF. RT-PCR from isolated fat body and abdominal integuments, together with in situ hybridization performed on puparia at this stage, showed that atf3 mRNA was primarily present in the larval epidermis (LECs) during the expression peak at 6 hours APF. From the time of head eversion (12 hours APF) the mRNA level remained low until the second day of pupal development, and then it grew steadily during morphogenesis of the adult. Quantitative RT-PCR revealed a 4.3-fold difference in atf3 mRNA abundance between 0 and 72 hours APF. In contrast to the tight regulation of atf3, the mRNAs of fos and jun fluctuated little during the examined period. Therefore, unlike Jun or Fos, Atf3 was dynamically regulated during metamorphosis at the level of transcription (Sekyrova, 2010).
The precise temporal control of atf3 expression suggested that the rise and subsequent fall of Atf3 during metamorphosis might be critical for the complex morphogenesis occurring in fly pupae. This possibility was tested by means of sustained expression of the full-length Atf3 protein using the UAS-Gal4 system with various drivers. A striking, fully penetrant metamorphic defect was observed with the pumpless (ppl) Gal4 driver. Although ppl>atf3 animals developed normally until the pupal stage, they failed to complete fusion of the adult abdominal epidermis. A dorsal cleft in the abdomen remained that could not be covered with the adult cuticle, and consequently 86% of the flies died inside the puparium. All of the ppl>atf3 adults that did eclose showed abdominal lesions filled with the old pupal cuticle lacking adult pigmentation and bristles, often with a clot covering a bleeding wound. Adults with the same abdominal cleft (but otherwise normal) also emerged when atf3 was moderately and ubiquitously misexpressed under the arm-Gal4 driver, suggesting that abdominal morphogenesis was the process most sensitive to ectopic Atf3 (Sekyrova, 2010).
The adult fly abdomen derives from histoblasts that proliferate, replace LECs and finally differentiate, giving rise to the adult cuticle. Therefore, the observed abdominal defect suggested a compromised function of the epidermis, either LECs, histoblasts or both cell types. To distinguish between these possibilities, expression of the ppl-Gal4 driver was first examined in the epidermis. It was found that ppl-Gal4 was active in LECs but not in histoblasts. Second, another driver, Eip71CD-Gal4, which was inactive in histoblasts but strongly expressed in LECs, was examined. Eip71CD-Gal4-driven misexpression of atf3 mostly produced lethal pupae lacking adult cuticle, but it occasionally yielded adults with a dorsal abdominal cleft. In addition to being active in LECs, both ppl-Gal4 and Eip71CD-Gal4 (data not shown) were also expressed in the fat body. However, no abdominal defects occurred when atf3 was misexpressed under either of three fat-body-specific Gal4 drivers, Lsp2, Cg or C7. Third, to rule out the possibility that ectopic Atf3 affected the imaginal epidermis, its expression was directed to histoblasts by using the escargot (esg) and T155 Gal4 drivers; in neither case the fusion of the adult abdominal epidermis was affected (Sekyrova, 2010).
To finally confirm that abdominal morphogenesis was disrupted by sustained atf3 activity in LECs, atf3 was induced by using the flp-out technique. Owing to the timing of heat-shock induction to the mid-third instar, this method triggers expression in the polyploid larval cells but not in the diploid histoblasts . Misexpression of atf3 under the actin promoter following the flp-out event invariantly led to an abdominal cleft. The lesions were often more severe than those observed in ppl>atf3 animals, affecting also lateral and ventral parts of the abdomen. Together, the above data demonstrate that the sustained expression of atf3 prevents fusion of the adult abdominal epidermis by acting upon LECs, suggesting that the replacement of these obsolete larval cells by adult histoblasts requires the developmental downregulation of atf3 expression (Sekyrova, 2010).
To understand the cellular events underlying the incomplete epithelial closure in ppl>atf3 animals, cell membranes were visualized by antibody staining of the septate junction component, Discs large 1 (Dlg1), or used a transgenic DE-cadherin::GFP fusion protein (shg::gfp). In wild-type animals 24 hours APF, LECs covering the surface of the abdomen gave way to the rapidly expanding nests of histoblasts that began to fuse laterally and ventrally. In ppl>atf3 pupae the histoblast nests also spread, and at least at 16 hours APF, before their fusion, they comprised normal numbers of histoblasts. By 48 hours APF a control abdomen was fully covered with adult epidermis consisting exclusively of histoblasts, now forming sensory bristles. Histoblasts in ppl>atf3 abdomens also differentiated the adult cuticle with sensory bristles, although polarity of the bristles near the dorsal cleft was altered. However, in contrast to the control, a large population of LECs remained in the dorsal abdomen of ppl>atf3 animals at 48 hours APF. The membranes of the persisting LECs accumulated the Dlg protein, and although these cells became severely deformed they survived throughout metamorphosis to the adult stage. When visualized in live ppl>atf3 pupae, the apical junctions of the remaining LECs displayed interdigitation and accumulation of DE-cadherin::GFP. Another adherens junction component, the Drosophila β-catenin Armadillo, was also enriched in atf3-expressing LECs (Sekyrova, 2010).
Cooperation between adherens junctions and the apical ring of actomyosin cytoskeleton is required for basal extrusion of LECs. The altered pattern of DE-cadherin and β-catenin therefore suggests that excessive Atf3 might prevent LEC extrusion through stabilization of the cell-cell adhesion complex. To examine the effect of Atf3 on LECs in further detail, the flp-out technique, which allows comparisons of atf3-misexpressing and control LECs within one tissue, was employed. Membrane interdigitation occurred between atf3-positive LECs already at 18 and 24 hours APF, even in areas where the LECs had no contact with histoblasts. At 48 hours APF only LECs expressing atf3 persisted, apparently being squeezed by the expanding histoblasts. The membrane-associated DE-cadherin::GFP signal was stronger in adjacent atf3-positive LECs compared with non-induced LECs, and quantitative analysis of confocal images acquired at 18 hours APF and at 24 hours APF both revealed a statistically significant 1.4-fold increase of the DE-cadherin::GFP signal intensity upon atf3 induction. Enrichment of DE-cadherin on apical membranes of atf3-expressing LECs was further confirmed on confocal cross sections (Sekyrova, 2010).
Although some atf3-positive LECs began the extrusion process, they could not detach from the apical surface even when entirely surrounded by histoblasts, possibly being tethered to it by the excessive adhesion protein. By contrast, control LECs did completely separate from the epithelium. In addition, LECs overexpressing atf3 displayed apical enrichment of moesin, an actin-binding protein of the ERM (ezrin, radixin, moesin) family, which links transmembrane proteins to cortical actin filaments. Interestingly, prominent accumulation of DE-cadherin was also observed in atf3-expressing clones of epithelial cells within the hinge region of wing discs that form the adult thorax, indicating that the effect of Atf3 on cell adhesion components may not be limited to larval epithelia (Sekyrova, 2010).
In summary, these results show that deregulation of atf3 expression causes marked changes of cell membranes, including interdigitation and accumulation of cell adhesion molecules, suggesting that LEC adhesiveness might be increased. Although some of the affected LECs initiate extrusion, this process stays incomplete. Consequently, the adhering LECs present a physical barrier for the migrating histoblasts (Sekyrova, 2010).
Rho kinase (Rok)-dependent phosphorylation of myosin regulatory light chain was shown to be required for LEC extrusion. To examine a possible relationship between the Rok-dependent cytoskeletal regulation and Atf3, the function of the GTPase Rho1 (also called RhoA), which acts immediately upstream of Rok, was disrupted. RNAi silencing of Rho1 using the ppl-Gal4 driver produced a phenocopy of atf3 misexpression, causing a dorsal abdominal cleft in 100% of ppl>Rho1(RNAi) adults, of which most died in the puparium and about 12% eclosed, similar to ppl>atf3 animals. However, when Rho1 RNAi and misexpression of atf3 in LECs were combined, the abdominal defect became more severe, not allowing any pharate adults to eclose. Conversely, co-expression of a dominantly active Rho1V14 protein suppressed the otherwise fully penetrant abdominal defect in some ppl-atf3 flies. Surprisingly, it was found that the endogenous Rho1 protein was mislocalized in atf3-misexpressing LECs, showing a diffuse cytoplasmic signal, compared with membrane localization in control LECs. These results suggest a genetic interaction between Rho signaling and atf3, and support the idea that excess Atf3 prevents extrusion of LECs by altering their cell adhesion properties (Sekyrova, 2010).
Disturbed function of the ecdysone receptor (EcR) has been shown to prevent extrusion of LECs, causing a dorsal abdominal cleft that closely resembles the Atf3 gain-of-function phenotype. Therefore whether stimulating EcR-dependent signaling by addition of the natural agonist 20E might overcome the defect caused by sustained atf3 expression was examined. Indeed, supplying third-instar ppl>atf3 larvae with dietary 20E increased the number of eclosing adults, the abdominal scars of which were in 22% of the cases partially or completely sealed with normal adult cuticle (Sekyrova, 2010).
Atf3 interacts with Jun to form a DNA-binding complex and genetically when overexpressed in the developing compound eye. To see if this interaction is biologically relevant during abdominal morphogenesis, whether Atf3 relies on the presence of Jun to cause the dorsal cleft phenotype was tested. First, it was confirmed that Jun is indeed expressed in LECs during metamorphosis. RNAi-mediated depletion of Jun in animals that misexpressed atf3 under the ppl-Gal4 driver restored viability of adults from 14% (atf3 alone) to 100%. Strikingly, 87% of the ppl>atf3, jun(RNAi) adults eclosed with a completely normal abdomen. By contrast, RNAi knockdown of Fos in ppl>atf3 background did not improve the abdominal defect. RNAi silencing of either jun or fos alone under the ppl-Gal4 driver had no effect on the abdomen. These results demonstrate that Atf3 requires its partner Jun but not Fos to disrupt abdominal morphogenesis. Similar to the situation in the compound eye, the effect of misexpressed atf3 can be neutralized by simultaneously expressing Fos or its truncated bZIP domain under the ppl-Gal4 driver. Therefore, the model in which Atf3 and Fos compete for their common partner Jun may be extended to the developing abdomen (Sekyrova, 2010).
This study has identified Atf3 as a new partner of Jun in Drosophila. Previously, Jun has only been known to dimerize with itself and with the Drosophila homolog of Fos. Functional analysis of Atf3 has not yet been reported. These biochemical data show that, similar to mammalian ATF3 and JDP2, the Atf3 protein selectively binds Jun but not Fos. Also consistent with the properties of ATF3 and JDP2 is the ability of Atf3 to bind the ATF/CRE response element alone or synergistically with Jun. In contrast to its mammalian counterparts, however, neither Atf3 alone nor in complex with Jun bound to the AP-1 element under the same conditions. The selective interactions of Atf3 point to distinct biological roles for the Atf3-Jun and the Fos-Jun dimers, respectively (Sekyrova, 2010).
This study has shown a genetic interaction between Atf3 and Jun. The evidence is based on the ability of ectopic Atf3 to disturb morphogenesis of the adult abdomen and the compound eye, which strictly depends on the availability of Jun. Importantly, none of the Atf3 gain-of-function phenotypes could be induced by misexpression of the truncated bZIP domain of Atf3, suggesting that the functional Atf3 protein in complex with Jun is required. Based on the selectivity of Atf3 in a DNA-binding assay, it is predicted that the Atf3-Jun complex regulates specific target genes distinct from those targeted by Fos-Jun dimers (Sekyrova, 2010).
The data also reflect a relationship between the AP-1 and Atf3-Jun complexes. Although Fos does not dimerize or bind DNA with Atf3, its ability to suppress the Atf3 misexpression phenotype in the eye suggests that Fos and Atf3 compete in vivo for their common partner Jun. The fact that the same suppression can be achieved by overexpressing either the truncated Fos bZIP domain or Fos lacking phosphorylation sites indicates that the suppression does not rely on a transcriptional function of Fos but probably occurs through sequestering of Jun, even by a transcriptionally inactive Fos protein. Early in vitro studies have proposed a competition model for the AP-1 and Atf3 proteins to explain a temporal regulation of gene expression in the regenerating liver. However, to date such a relationship among Fos, Jun and Atf3 has not been supported with direct genetic evidence (Sekyrova, 2010).
Removal of LECs is normally complete by 36 hours APF, at which time the sheets of histoblasts reach the dorsal midline. The data strongly support the argument that the temporal downregulation of atf3 expression during abdominal morphogenesis is necessary for LECs to be replaced by the adult epidermis. When experimentally sustained, atf3 activity in LECs interfered with this exchange by blocking extrusion and death of the LECs. This was evident as the atf3-expressing LECs survived within the epithelial layer for days after their scheduled destruction (Sekyrova, 2010).
Interdigitation of cell membranes and accumulation of adherens junction proteins in LECs suggested that ectopic Atf3 caused adjacent LECs to reinforce their mutual contacts. This probably resulted from altered distribution of the proteins, as levels of the shg (DE-cadherin) mRNA remained unchanged in LECs of ppl>atf3 animals. By contrast, junctions between atf3-expressing LECs and their normal neighbors or histoblasts were smooth and presumably less rigid. DE-cadherin was similarly enriched in clones of imaginal disc cells. These observations suggested that differential adhesion of atf3-expressing cells might have led to their sorting out from the surrounding epithelium. Even modest differences in cadherin levels have been shown to cause segregation of cells within a population by altering their adhesiveness (Sekyrova, 2010).
Recent live imaging data have revealed that migrating histoblasts push the LECs ahead of themselves towards the dorsal midline, where histoblasts fuse last. The atf3-expressing LECs that adhered to each other were probably moved and pressed by the expanding histoblasts to the dorsal side, whereas non-induced LECs were eliminated. This explains why the abdominal lesions primarily occurred at the dorsal midline, although flp-out experiments showed that atf3 misexpression could affect LECs in other areas as well. Strengthened contacts among persisting LECs probably blocked invasion of histoblasts in between them and inhibited LEC extrusion, eventually causing gaps in the adult epidermis (Sekyrova, 2010).
In accord with the notion that extrusion from the epithelium is a prerequisite for LECs to undergo apoptosis, it is assumed that sustained presence of Atf3 primarily enhanced adhesiveness of LECs, which only consequently prevented their death. This view is supported by the observation that membranes of atf3-expressing LECs interdigitated and accumulated DE-cadherin as early as 18-24 hours APF, even in areas of the larval epidermis that were far from histoblasts and where control LECs did not yet extrude. In addition, the Atf3 gain-of-function phenotype was stronger than abdominal closure defects caused by caspase mutation or inhibition. When the anti-apoptotic proteins p35 or DIAP1 (Thread FlyBase) was misexpressed under the ppl-Gal4 driver, the resulting dorsal lesions were not lethal and were clearly milder than the broad, mostly fatal scars in ppl>atf3 animals. Compared with the large contiguous populations of persisting LECs in ppl>atf3 pupae, inhibiting apoptosis with p35 only allowed small islands of LECs to survive (Sekyrova, 2010).
Ecdysone signaling promotes replacement of the abdominal epithelia by stimulating both the early histoblast proliferation and the extrusion of LECs. As atf3 misexpression affected LECs but did not impair early histoblast proliferation, the latter possibility remains, that added 20E counteracted the effect of ectopic Atf3 by facilitating the extrusion process. Since normal 20E titers was detected in ppl>atf3 larvae or prepupae, the failure of LEC extrusion was not a result of steroid deficiency. Also, 20E had no effect on atf3 mRNA levels, at least in Drosophila S2 cells or third-instar larvae. Atf3 and ecdysone signaling therefore probably influence LEC extrusion by acting independently (Sekyrova, 2010).
Although the mechanism through which ecdysone contributes to LEC removal is unknown, one attractive possibility is that it might cooperate with Rho signaling, which is required for LEC extrusion as well. It has been demonstrated that genetic interaction between the 20E-response gene broad and components of the Rho pathway including RhoGEF2, Rho1 and myosin II is important for ecdysone-dependent epithelial cell elongation during Drosophila leg morphogenesis. The current data show that Rho1 becomes mislocalized in LECs upon atf3 misexpression and that Rho1 silencing enhances the abdominal gain-of-function phenotype of atf3. The exact relationship between Atf3, Rho1 and ecdysone remains to be determined. However, Atf3 clearly represents a new intrinsic regulator of epithelial cell replacement during Drosophila metamorphosis (Sekyrova, 2010).
Juvenile hormone (JH) regulates many developmental and physiological events in insects, but its molecular mechanism remains conjectural. Genetic ablation of the corpus allatum cells of the Drosophila ring gland (the JH source) results in JH deficiency, pupal lethality and precocious and enhanced programmed cell death (PCD) of the larval fat body. In the fat body of the JH-deficient animals, Dronc and Drice, two caspase genes that are crucial for PCD induced by the molting hormone 20-hydroxyecdysone (20E), are significantly upregulated. These results demonstrated that JH antagonizes 20E-induced PCD by restricting the mRNA levels of Dronc and Drice. The antagonizing effect of JH on 20E-induced PCD in the fat body was further confirmed in the JH-deficient animals by 20E treatment and RNA interference of the 20E receptor EcR. Moreover, MET and GCE, the bHLH-PAS transcription factors involved in JH action, were shown to induce PCD by upregulating Dronc and Drice. In the Met- and gce-deficient animals, Dronc and Drice were downregulated, whereas in the Met-overexpression fat body, Dronc and Drice were significantly upregulated leading to precocious and enhanced PCD, and this upregulation could be suppressed by application of the JH agonist methoprene. For the first time, this study demonstrates that JH counteracts MET and GCE to prevent caspase-dependent PCD in controlling fat body remodeling and larval-pupal metamorphosis in Drosophila (Liu, 2009).
The status quo action of JH has been well documented in several insect orders, particularly in Coleoptera, Orthoptera and Lepidoptera, in which JH treatment causes supernumerary larval molting and JH deficiency triggers precocious metamorphosis. However, as JH does not cause supernumerary larval molting in flies, evidence for the status quo action of JH in Drosophila has remained elusive. From past studies and from the experimental data presented in this study, it is concluded that the status quo hypothesis does indeed apply to JH action in Drosophila. First, although JH application during the final larval instar or during the prepupal stage has little effect on the differentiation of adult head and thoracic epidermis in Drosophila, it does prevent normal adult differentiation of the abdominal epidermis. After JH treatment, a second pupal, rather than an adult, abdominal cuticle is formed in Diptera. Second, JH or a JH agonist applied to Drosophila at the onset of metamorphosis results in lethality during pupal-adult metamorphosis. Similarly, global overexpression of jhamt (Juvenile hormone acid methyl transferase) results in severe defects during the pupal-adult transition and eventually death (Niwa, 2008). Third, CA ablation leading to JH deficiency causes precocious and enhanced fat body PCD. Fourth, JH deficiency results in pupal lethality and delayed larval development, although JH deficiency is not sufficient to cause precocious metamorphosis. The composite data demonstrate that JH in Drosophila does have status quo actions on the abdominal epidermis during pupal-adult metamorphosis and on the fat body during larval-pupal metamorphosis. It is concluded that the status quo action of JH in Drosophila is functionally important, but more subtle than that in Coleoptera, Orthoptera and Lepidoptera. However, it is not clear whether JH is essential for embryonic and earlier larval development because the CA cells are not completely ablated in the JH-deficient animals until the early-wandering (EW) stage. To address this question, it would be necessary to generate a mutant (i.e., of jhamt) that interrupts JH but not the farnesyl pyrophosphate biosynthesis pathway (Liu, 2009).
The insect fat body is analogous to vertebrate adipose tissue and liver and
functions as a major organ for nutrient storage and energy metabolism. In response to 20E pulses, Drosophila larval organs undergo a developmental
remodeling process during metamorphosis. Blocking
the 20E signal specifically in the fat body during the larval-pupal transition
(Lsp2>; UAS-EcRDN) prevented the fat body from
undergoing PCD and cell dissociation (Liu, 2009).
The experimental data in this paper demonstrates that JH prevents
caspase-dependent PCD in the fat body during the larval-pupal transition in
Drosophila. First, JH deficiency in Aug21>, UAS-grim
resulted in the fat body undergoing precocious and enhanced PCD and cell
dissociation. Aug21> is a GAL4 driver that specifically targets gene expression to the CA. Precocious and enhanced apoptosis appeared as early as L3D1 in the JH-deficient animals. Methoprene application on L3D1 was able to rescue ~40% of the pupae to adults, but it failed to rescue post-EW. Second, 2D-DIGE/MS
and qPCR analyses indicated that the fat body in the JH-deficient animals has
multiple developmental defects. The upregulation of the caspase genes
Dronc and Drice should account for the PCD in the fat body, as
overexpression of Dronc in the fat body causes PCD, cell
dissociation, and thus lethality. Overexpression of Dronc or Drice
in cells and tissues is sufficient to cause caspase-dependent PCD. Third, the
20E-triggered transcriptional cascade in the fat body was downregulated in the
JH-deficient animals, indicating that JH does not suppress the 20E-triggered transcriptional cascade in preventing caspase-dependent PCD in the fat body (Liu, 2009).
The antagonizing effect of JH on 20E-induced PCD in the fat body was
further confirmed in the JH-deficient animals by 20E treatment and RNA
interference of EcR. One might expect that perfect timing, titer and receptor response of JH and 20E are required to ensure accurate PCD in a tissue- and stage-specific manner during Drosophila metamorphosis. In the
JH-deficient animals, the upregulation of Dronc and Drice
resulted in precocious and enhanced PCD, such that the JH-deficient animals
are committed to die during the larval-pupal transition. This hypothesis was
strengthened by overexpression of Dronc specifically in the fat body,
which caused larval lethality. Taken together, it is concluded that JH antagonizes 20E-induced caspase-dependent PCD in controlling fat body remodeling and larval-pupal metamorphosis in Drosophila (Liu, 2009).
Based on the phenotypes and gene expression profiles in the four fly lines
used, it is concluded that JH counteracts MET and GCE to prevent caspase-dependent PCD. First, the Met-overexpressing animals died during larval life, with precocious and enhanced PCD and cell dissociation in the fat body.
Dramatic upregulation of Dronc and Drice was observed when
Met was specifically overexpressed in the fat body and this
upregulation was significantly decreased by methoprene application demonstrating that JH is epistatic to MET and GCE. Moreover, the Dronc-overexpressing
animals exhibited similar phenotypes to the Met-overexpressing animals. Second, in the fat body of the JH-deficient animals, PCD and the expression of
Dronc and Drice were upregulated but not as significantly as in the
Met-overexpressing animals. This might explain why the JH-deficient
animals did not die until early pupal life. Third, both the
global JH-overexpressing animals and the Met/gce-deficient animals
died during the pupal-adult transition. In these
animals, Dronc and Drice were downregulated and
caspase-dependent PCD was decreased in the fat body, implying that these animals died from a lack of caspase-dependent PCD. Weak mutants of Dronc and Drice mutants die during
pupal life, showing that caspase-dependent PCD is essential for Drosophila metamorphosis. In addition, it was also observed that methoprene application at the onset of metamorphosis results in delayed fat body remodeling (Liu, 2009).
In the future, it will be crucial to elucidate the detailed molecular
mechanism of how JH counteracts MET and GCE to prevent caspase-dependent PCD.
In Drosophila S2 cells, the transcriptional activity of MET is
dependent on the JH concentration and both MET-MET and MET-GCE interactions can be greatly diminished by JH. The bHLH-PAS transcription factors typically function as hetero- or homodimers. If MET/GCE is the Juvenile Hormone Receptor (JHR), the transcriptional activities of
the dimerized MET/GCE and the JH-MET/GCE complex should differ. In other
words, the dimerized MET/GCE should induce transcription of Dronc and
Drice and, in turn, JH binding to form the JH-MET/GCE complex should
reduce this induction. Although there are no examples in
the literature in which a receptor, without ligand, acts as a transcriptional
activator and the transcriptional activity of the receptor is diminished when
the ligand is bound, it could be speculated that the JHR is a unique hormone
receptor and perhaps that is the reason why it has yet to be isolated and
characterized. Unfortunately, the experiments described here were
conducted in Drosophila S2 cells, where the possibility of an
endogenous JHR could not be eliminated. Although MET/GCE is definitely a key
component in the JH signal transduction pathway, whether MET/GCE is the bona
fide JHR remains conjecture (Liu, 2009).
It is very likely that MET cross-talks with EcR-USP via a large molecular
complex. One can hypothesize that MET promotes 20E action in the absence of JH and suppresses 20E action in the presence of JH, a model which is favored.
Drosophila FKBP39 (FK506-BP1) could be a key component in this
complex because it physically interacts with MET, EcR and USP, and binds the
D. melanogaster JH response element 1. Moreover, Drosophila FKBP39 inhibits 20E-induced autophagy (Juhász., 2007).
Further analysis of the complex will be crucial to precisely define the
molecular mechanism of cross-talk between the action of JH and 20E (Liu, 2009).
In summary, it is concluded that JH counteracts MET and GCE to prevent
caspase-dependent PCD in controlling fat body remodeling and larval-pupal
metamorphosis in Drosophila. The Drosophila fat body has provided an excellent model for studying the long-standing question of JH signal transduction. To finally settle the question of the bona fide JHR and to understand the precisely defined molecular mechanism of JH action requires
further research at a variety of levels in several species of insects that can be genetically manipulated, such as Drosophila, Bombyx and Tribolium (Liu, 2009).
Juvenile hormone (JH) biosynthesis in the corpus allatum (CA) is regulated by neuropeptides and neurotransmitters produced in the brain. However, little is known about how these neural signals induce changes in JH biosynthesis. This study reports a novel function of TGFβ signaling in transferring brain signals into transcriptional changes of JH acid methyltransferase (jhamt), a key regulatory enzyme of JH biosynthesis. A Drosophila genetic screen identified that Tkv and Mad are required for JH-mediated suppression of broad (br) expression in young larvae. Further investigation demonstrated that TGFβ signaling stimulates JH biosynthesis by upregulating jhamt expression. Moreover, dpp hypomorphic mutants also induces precocious br expression. The pupal lethality of these dpp mutants is partially rescued by an exogenous JH agonist. Finally, dpp is specifically expressed in the CA cells of ring glands, and its expression profile in the CA correlates with that of jhamt and matched JH levels in the hemolymph. Reduced dpp expression was detected in larvae mutant for Nmdar1, a CA-expressed glutamate receptor. Taken together, it is concluded that the neurotransmitter glutamate promotes dpp expression in the CA, which stimulates JH biosynthesis through Tkv and Mad by upregulating jhamt transcription at the early larval stages to prevent premature metamorphosis (Huang, 2011).
The functions of the TGFβ superfamily and other morphogens in regulating insect metamorphosis are rarely reported. In two independent genetic screens, it was discovered that Drosophila TGFβ signaling controls two different aspects of insect metamorphosis. In a previous study, it was found that Baboon (Babo) and dSmad2-mediated TGFβ signaling regulates larval neuron remodeling, which is part of the insect central nervous system metamorphosis induced by 20E during the pupal stage. Further investigation revealed that Babo/dSmad2-mediated TGFβ signaling controls larval neuron remodeling through regulating the expression of EcR-B1, a specific isoform of the 20E receptor (Huang, 2011).
This paper reports several findings. First, br is precociously expressed in 2nd instar tkv and Mad mutant larvae. Second, the precocious br expression phenotype in tkv and Mad mutant larvae can be suppressed by exogenous JH agonist (JHA). Third, Tkv and Mad repressed br expression in a non-cell-autonomous manner. Fourth, the presence of Mad in the CA is sufficient to repress br expression in the fat body (FB). Fifth, jhamt mRNA levels and JHAMT activity were significantly reduced in the Mad-deficient larvae. These results demonstrate that Tkv- and Mad-mediated signaling is required in the CA to activate jhamt expression and thus JH biosynthesis, which in turn controls insect metamorphosis (Huang, 2011).
The Drosophila genome encodes two TGFβ type II receptors, Punt (Put) and Wishful thinking (Wit). The genetic screen failed to identify a role for either of these receptors in the regulation of JH biosynthesis. Put and Wit are most probably functionally redundant in this biological event, as in the case of TGFβ-mediated mushroom body neuron remodeling (Huang, 2011).
Dpp is a key morphogen that controls dorsal/ventral polarity, segmental compartment determination and imaginal disc patterning. Dpp function usually depends on its gradient distribution. In an attempt to identify the ligand for Tkv/Mad-mediated TGFβ signaling in the CA, a novel, gradient-independent role for Dpp was discovered that controls JH biosynthesis. Dpp is the ligand of Tkv, which regulates jhamt transcription. Loss of Dpp, even RNAi reduction of Dpp in the CA specifically, causes precocious br expression at the early larval stages, which phenocopies tkv and Mad mutants. Phenotypes of dpp, including precocious br expression and lethality, are at least partially rescued by JHA treatment or ectopic jhamt expression in the CA. Notably, dpp-lacZ is strictly expressed in the CA cells, but not in the other two types of endocrine cells in the ring gland: the prothoracic gland and corpus cardiacum cells. The developmental expression profile of dpp in the CA is always consistent with that of jhamt. Finally, dpp expression in the CA may be directly controlled by neurotransmitter signals in the brain, which is supported by reduced dpp and jhamt transcription levels in the Nmdar1 mutant wandering larvae (Huang, 2011).
Several lines of evidence suggest that Met is a crucial regulator at or near the top of a JH signaling hierarchy, possibly acting as a JH receptor. However, null Met mutants of Drosophila are completely viable, which is unexpected if Met is a JH receptor. A recent investigation indicated that another Drosophila bHLH-PAS protein, Germ cell-expressed (Gce), which has more than 50% homology to Met, may function redundantly to Met in transducing JH signaling (Baumann, 2010). Because Met is on the X chromosome in the fly genome, it was not covered by the genetic screen. The br protein in the FBs of a Met null allele, Met27, was tested at the 2nd instar larval stage, and precocious br expression was observed. Importantly, this precocious br expression phenotype could not be suppressed by exogenous JHA. This result not only supports the previous reports regarding the function of Met in transducing JH signaling but also suggests that the precocious br expression is a more sensitive indicator for the reduced JH activity in Drosophila compared with precocious metamorphosis, lethality and other phenotypes (Huang, 2011).
Kr-h1 was reported to act downstream of Met in mediating JH action. Studies in both Drosophila and Tribolium reveal that, at the pupal stages, exogenous JHA induces Kr-h1 expression, which in turn upregulates br expression. The genetic screen successfully identified that Kr-h1 is cell-autonomously required for the suppression of br expression at young larval stages. Precocious br expression occurred in the FBs of Kr-h1 mutants and was not suppressed by JHA treatment. Therefore, these studies further suggest that Kr-h1 functions as a JH signaling component in mediating insect metamorphosis. However, the finding shows that, at the larval stages of Drosophila, the JH-induced Kr-h1 suppresses, rather than stimulates, br expression. This result is consistent with the facts that Kr-h1 functions to prevent Tribolium metamorphosis and Br is a crucial factor in promoting pupa formation (Huang, 2011).
In summary, this study has found a novel function of Dpp, Tkv and Mad-mediated TGFβ signaling in controlling insect metamorphosis. As summarized in a model, the brain sends neurotransmitters, such as glutamate, to the CA through neuronal axons. Glutamate interacts with its receptor (NMDAR) on the surface of CA cells to induce dpp expression. Dpp protein produced and secreted by CA cells forms a complex with TGFβ type I receptor (Tkv) and type II receptor on the membrane of CA cells, followed by phosphorylation and activation of Tkv. Activated Tkv in turn phosphorylates Mad, which is imported into the nucleus together with co-Smad and stimulates jhamt expression. JHAMT in CA cells transforms JH acid into JH, which is released into hemolymph. The presence of JH in young larvae prevents premature metamorphosis through Met/Gce and Kr-h1 by suppressing the expression of br, a crucial gene in initiating insect metamorphosis (Huang, 2011).
During the development of multicellular organisms, body growth is controlled at the scale of the organism by the activity of long-range signaling molecules, mostly hormones. These systemic factors coordinate growth between developing tissues and act as relays to adjust body growth in response to environmental changes. In target organs, long-range signals act in concert with tissue-autonomous ones to regulate the final size of a given tissue. In Drosophila, the steroid hormone ecdysone plays a dual role: peaks of secretion promote developmental transitions and maturation, while basal production negatively controls the speed of growth. The antagonistic action of ecdysone and the conserved insulin/insulin growth factor (IGF) signaling pathway regulate systemic growth and modulate final body size. This study has unraveled an unexpected role of bantam microRNA in controlling body size in Drosophila. The data reveals that, in addition to its well-characterized function in autonomously inducing tissue growth, bantam activity in ecdysone-producing cells promotes systemic growth by repressing ecdysone release. Evidence is provided that the regulation of ecdysone production by insulin signaling relies on the repression of bantam activity. These results identify a molecular mechanism that underlies the crosstalk between these two hormones and add a new layer of complexity to the well-characterized role of bantam in growth control (Boulan, 2013).
Because ban and ecdysone affect systemic growth in an
opposite manner, it is likely that ban acts in PG cells by preventing
ecdysone production. The circulating
levels of the active form of ecdysone (20E) were measured in in P0206>ban
animals at two developmental time points: at the beginning
of the wandering phase (early L3w), and just before the larva/
pupa transition (late L3w). To do so,
wandering larvae were precisely staged by monitoring gut clearance of blue food. As expected, 20E levels were already high in early L3w (blue gut) control larvae and further peaked in late L3w (clear
gut). In contrast, P0206>ban larvae showed lower circulating
20E levels than controls at both stages, and the amplitude
of the peak was strongly reduced. Ecdysone
signaling in target tissues was also reduced in late larval development
upon ban overexpression in the PG (in phm>banD
animals) as measured by the expression levels of
E75A and Broad-Complex (BR-C), two targets of EcR. Furthermore, the phantom (phm), disembodied (dib), and
shade (sad) genes, which are specifically expressed in the
PG and encode enzymes required for ecdysteroid biosynthesis, showed reduced expression in
phm>ban animals. Consistent with the reduced
levels of 20E production and signaling in phm>ban larvae, an
increase in 20E levels, produced by feeding the animals ecdysone-
supplemented medium, rescued pupa formation. These findings indicate that targeted expression of
ban in ecdysone-producing cells has a negative impact on
20E production (Boulan, 2013).
banΔ1 mutant larvae displayed high lethality in late larval
development. Thus, ecdysone signaling was assessed
in these larvae during earlier stages. Two different
developmental points precisely staged with respect to the
transition from second (L2) to third (L3) larval instar were
selected: 2 hr and 20 hr after ecdysis to the third instar (AL3E).
No changes in the expression of BR-C were detected, most probably because at that time, 20E levels had
not yet reached the minimum threshold to activate this target.
However, the quantification of E75A mRNA levels revealed
higher expression in banΔ1 larvae when compared to controls, as did the quantification of dib, phm, and sad mRNA levels. Ni predicted ban
target site was found in the 3' UTR of these genes, suggesting
that this repression is not direct (Boulan, 2013).
Ecdysone signaling negatively regulates body growth. To
test whether the undergrowth phenotype observed in ban
mutants is a result of abnormally high ecdysone levels, whole-mount ecdysone signaling was reduced by removing one copy of EcR or impaired ecdysone synthesis by
depleting the levels of Sad and Phm in the PG
of banΔ1 animals. In all cases, the size of banΔ1 pupae was
largely rescued. Collectively, these results suggest that ban
participates in reducing ecdysone production in PG cells and
corroborate the hypothesis that the systemic growth defects
observed in ban mutants are caused by increased ecdysone
levels. Consistent with the cell-autonomous growth-promoting
role of ban, the PG was larger in P0206>ban
animals (that produce less ecdysone) than in controls,
whereas it was much smaller in ban mutants.
Thus, the impact of ban on ecdysone production is not a
consequence of changes in PG size (Boulan, 2013).
Despite displaying higher levels of ecdysone, banΔ1 mutant
animals reached metamorphosis with a delay. It
has been reported that a strong reduction in larval growth rates
can affect developmental timing as a result of a delay in the
attainment of critical size for metamorphosis. In order to
address whether banΔ1 animals are delayed as a consequence
of their reduced growth rates, as
a simple proxy of critical size, the time at which the minimal
viable size for metamorphosis was achieved in was determined banΔ1 mutant
and wild-type animals. Larvae were synchronized at the
second (L2) to third (L3) instar transition and then starved at
fixed time points to assess survival and capacity to enter
into metamorphosis. Remarkably, banΔ1 larvae reached the
threshold of 50% of survival with a delay when compared to
wild-type animals. These data, together with the
fact that targeted expression of ban in the ring gland largely
rescued growth rates and developmental delay of banΔ1
animals, support the proposal that the
developmental delay is at least in part a consequence of
reduced growth rates. Other activities of ban, such as reduced
growth of the imaginal tissues or impaired dendrite development,
might also affect the timing of metamorphosis (Boulan, 2013).
The production of ecdysone is tightly controlled during larval development.
Under normal conditions, ecdysone levels are low during the growth period,
thereby allowing optimal body growth rates, and peak at the end of the third-instar
larval stage to induce entry into metamorphosis. To monitor whether
ban activity levels are also dynamically regulated in the PG, use was made of
a ban sensor that expresses GFP under control of a ubiquitously active tubulin
promoter and carries two perfect ban fixation sites in its 3' UTR thus making it
repressed in the presence of the miRNA. A control sensor lacking
the fixation sites showed high GFP expression both in early and late larval
PGs. ban sensor levels, however, were low in the PG of
second- and early third-instar larvae and considerably increased
in wandering third-instar larvae. This observation leads to the proposal that high
ban activity in young larvae contributes to the maintenance
of low ecdysone titers and the promotion of systemic growth,
whereas reduced activity in late PGs contributes to the generation
of the ecdysone peak, the cessation of growth, and entry into metamorphosis (Boulan, 2013).
What is the upstream signal that regulates ban activity?
The conserved insulin/insulin growth factor (IGF) signaling
pathway directly promotes growth in
target tissues and is the main relay to couple body growth to
nutritional state. In young feeding larvae, insulin signaling
in the PG also promotes the basal production of ecdysone, which in turn inhibits body growth. This buffering mechanism, based on the antagonistic action of insulin and
ecdysone, modulates final body size in response to nutritional
changes. Interestingly, ban activity levels were strongly reduced
in early PGs expressing different transgenes that activate
the insulin pathway. Increased levels of circulating Dilp2 also reduced ban activity in early PGs, as monitored by increased expression of the ban sensor. Thus, insulin signaling represses ban activity in ecdysone-producing cells (Boulan, 2013).
Thanks to a nutrient-sensing mechanism in the fat body, the equivalent to the vertebrate liver, food conditions control the secretion or expression of brain-derived Dilps, which are the main systemic supply of this hormone during the growth period. Consistent with the repression
caused by increased insulin signaling, young feeding
larvae growing on amino acid-rich medium showed a clear
decrease in ban activity in PG cells. This reduction depended on the enhanced activity
of Dilps, because the inhibition of insulin signaling in the PG
was sufficient to restore ban activity to normal levels (Boulan, 2013).
In order to address whether ban mediates the action of
insulin in regulating ecdysone production, genetic interactions were performed in gain- and loss-of-function conditions. Remarkably, the reduced body size phenotype obtained by
enhanced insulin signaling in the PG via several transgenes
was completely rescued by simultaneously increasing ban
levels in these cells. Given the fact that this rescue
implies a much greater effect on body size than the overexpression
of UAS-ban transgene alone, it was concluded that the
effects of ban and insulin signaling are not additive but rather
epistatic. This conclusion is further supported by the observation
that modulation of insulin signaling in the PG no longer
affected body size in a banΔ1 mutant background.
Altogether, these results indicate that the regulation of ecdysone
production by insulin signaling relies on the modulation
of ban activity in PG cells (Boulan, 2013).
So far, these results unravel a novel role of ban in promoting
larval body growth by reducing ecdysone production. In
contrast, ban was initially identified by its capacity to induce
organ growth in a cell-autonomous manner. That finding
prompted an exploration of the contribution of the systemic and
cell-autonomous activities of ban to organ growth. The imaginal
discs of Drosophila are epithelial sacs that grow in feeding
larvae to give rise after metamorphosis to the ectodermal
structures of the adult flies, such as legs, wings, or eyes. In
banΔ1 mutant larvae, the size of the wing imaginal discs
was strongly reduced when compared to control animals. Targeted expression
of ban in the PG partially rescued the wing growth defects
observed in ban mutant larvae. This result
supports the proposal that both the systemic and cell-autonomous
activities of ban are required to promote organ growth (Boulan, 2013).
In conclusion, these results establish that ban promotes systemic
growth by inhibiting the synthesis of the steroid
hormone ecdysone. During the growth period, ban mediates
the insulin-dependent regulation of ecdysone production
and therefore acts as a buffering mechanism to adjust final
body size in response to nutrient availability. Such a crosstalk
between insulin and steroid hormones and its impact on the
modulation of growth and developmental decisions are also
observed in Caenorhabditis elegans. Depending on environmental
conditions, the juvenile form of C. elegans either
enters maturation to give rise to an adult worm or arrests
development to form a dauer larva, a state that is specifically
adapted for survival. This decision is determined by the
levels of the steroid hormone dafachronic acid (DA). Mutations
that enhance insulin signaling, thereby mimicking a favorable
environment, increase DA levels and cause animals to
become incapable of forming dauer larvae. Remarkably,
the deletion of ban orthologs in C. elegans (the mir-58 family)
causes severe growth defects and prevents entry into the
dauer state under environmental stress (Alvarez-Saavedra, 2010). On the basis of
these observations, it is proposed that the role of ban in preventing
the production of steroid hormones in function of insulin
and nutrient levels might be conserved in other organisms in
order to regulate body growth and maturation (Boulan, 2013).
In Drosophila, growth takes place during the larval stages until the formation of the pupa. Starvation delays pupariation to allow prolonged feeding, ensuring that the animal reaches an appropriate size to form a fertile adult. Pupariation is induced by a peak of the steroid hormone ecdysone produced by the prothoracic gland (PG) after larvae have reached a certain body mass. Local downregulation of the insulin/insulin-like growth factor signaling (IIS) activity in the PG interferes with ecdysone production, indicating that IIS activity in the PG couples the nutritional state to development. However, the underlying mechanism is not well understood. This study shows that the secreted Imaginal morphogenesis protein-Late 2 (Imp-L2 - FlyBase name: Ecdysone-inducible gene L2), a growth inhibitor in Drosophila, is involved in this process. Imp-L2 inhibits the activity of the Drosophila insulin-like peptides by direct binding and is expressed by specific cells in the brain, the ring gland, the gut and the fat body. Imp-L2 is required to regulate and adapt developmental timing to nutritional conditions by regulating IIS activity in the PG. Increasing Imp-L2 expression at its endogenous sites using an Imp-L2-Gal4 driver delays pupariation, while Imp-L2 mutants exhibit a slight acceleration of development. These effects are strongly enhanced by starvation and are accompanied by massive alterations of ecdysone production resulting most likely from increased Imp-L2 production by neurons directly contacting the PG and not from elevated Imp-L2 levels in the hemolymph. Taken together these results suggest that Imp-L2-expressing neurons sense the nutritional state of Drosophila larvae and coordinate dietary information and ecdysone production to adjust developmental timing under starvation conditions (Sarraf-Zadeh, 2013).
In higher organisms, the duration of the juvenile stage needs to
be variable to ensure the development of a healthy and fertile
adult. Environmental stresses, such as adverse nutritional conditions, can delay development until a critical weight is reached.
Additional checkpoints ensure that increased growth rates,
induced by ideal nutritional conditions, do not lead to a premature
passage to the adult stage. In Drosophila, the juvenile growth stage
is terminated by pupae formation at the end of the third larval
instar. Larval/pupal transition is induced by a pulse of the steroid
hormone ecdysone produced by the PG (Sarraf-Zadeh, 2013).
Genetic manipulations of the Drosophila PG revealed the requirements of the IIS, Target of Rapamycin (TOR) and PTTH pathways to
control ecdysone production . Recently, IIS
dependent growth of the PG has been identified as an additional
factor controlling ecdysone production. Overexpression of PI3K, a
positive regulator of IIS, leads to premature, increased ecdysone
production resulting in a shortened L3 stage and early pupariation.
By contrast, overexpression of negative regulators of IIS in the PG
delays pupariation caused by lowered and delayed ecdysone
production. Reduction of whole organism IIS activity does not change critical weight but
delays its attainment. In contrast, ablation
of PTTH neurons induces a severe shift in critical weight, suggesting that these neurons play an important role in setting this
parameter. When larvae reach the critical
weight, PTTH is released on the PG and induces transcription of
genes involved in ecdysone production. However, PTTH expression is not modified
upon nutritional restriction, indicating that PTTH signaling does
not mediate starvation induced developmental delay. Signaling via TOR, the downstream kinase of IIS, links nutritional information to ecdysone production, since starvation induced developmental delay can
partially be rescued by upregulating TOR activity in the PG. This
suggests that downregulating TOR signaling upon starvation
desensitizes the PG for PTTH signals, resulting in delayed ecdysone
production. The present study shows
that increased IIS activity in the PG due to Imp L2 LOF rescues the
delay caused by malnutrition to a large extent, indicating that low
IIS also renders the PG irresponsive to the PTTH signal. Whether
the effects of low IIS in the PG are mediated by TOR or whether the
two pathways act independently remains to be elucidated (Sarraf-Zadeh, 2013).
Evidence is presented for a number of Imp L2 expressing
neurons to act as possible regulators of IIS activity in the PG. High
Imp L2 levels in the hemolymph can be excluded as possible
inhibitors of IIS signaling in the PG, since increasing hemolymph
levels of Imp L2 failed to reduce size and IIS activity of PG cells, but
resulted in a strong size decrease of the whole organism. On the
other hand, increasing Imp L2 levels in Imp L2 positive neurons
targeting the PG causes a massive decrease in PG size and lowers
IIS activity within PG cells. These results support the idea that the
PG does not receive information about the nutritional state of the
organism through the hemolymph but rather from Imp L2 expressing
neurons. Thus, this work reveals a novel local function of the
negative growth regulator Imp L2 in controlling IIS activity and
ecdysone production in the PG. This finding reveals a novel
mechanism for the spatial regulation of IIS: through locally
restricted effects of Imp L2, diverse tissues can be effectively
subjected to different levels of IIS (Sarraf-Zadeh, 2013).
Interestingly, the ability of IIS to coordinate growth with
development seems to be conserved throughout evolution. In
humans, the onset of puberty is linked to the nutritional state,
leading to early puberty in well fed western societies.
In contrast, juvenile females suffering from type I
diabetes mellitus display a notable delay in menarche,
indicating that decreased IIS also delays maturation in
humans. Moreover, in Caenorhabditis elegans, malnutrition during
the first larval stage leads to developmental arrest by inducing
dauer formation, which is a larval stage best adapted for survival
under adverse environmental conditions. Mutations reducing IIS pathway activity lead to dauer
formation independent of the nutritional state. Hence, different phyla developed similar strategies to cope
with adverse nutritional conditions during the juvenile state.
When IIS activity is below a certain threshold, development is
attenuated until sufficient nutrients are available, to ensure the
formation of healthy and fertile adults. In Drosophila larval malnutrition
leads to delayed pupariation, due to decreased IIS activity
in the PG which in turn delays the production of the steroid hormone ecdysone (Sarraf-Zadeh, 2013).
Steroid hormones also play an important role in human
development. In cases of human hypogonadism, puberty is prolonged, which can lead to abnormally tall adults if not treated with steroid substitutes. Referring the current data to the
human system, the putative Imp L2 homolog IGFBP 7 (also known
as IGFBP rP1) also displays a very diverse protein expression
pattern, indicating a specialized function in different organs. Amongst other tissues, IGFBP 7 is
expressed in different regions of the human brain, leading to the speculation that it might act as a local
regulator of steroid production as well (Sarraf-Zadeh, 2013).
In summary, the data provides novel insights into the coupling
of developmental cues to nutritional state. Since IIS and steroid
hormones play evolutionarily conserved roles in regulating growth
and development, the findings on the local function of the insulin
binding protein Imp L2 in controlling ecdysone production might
be of general interest (Sarraf-Zadeh, 2013).
EcR-dependent transcription, and thus, developmental timing in Drosophila, is regulated by CDK8 and its regulatory partner Cyclin C (CycC), and the level of CDK8 is affected by nutrient availability. cdk8 and cycC mutants resemble EcR mutants and EcR-target genes are systematically down-regulated in both mutants. Indeed, the ability of the EcR-Ultraspiracle (USP) heterodimer to bind to polytene chromosomes and the promoters of EcR target genes is also diminished. Mass spectrometry analysis of proteins that co-immunoprecipitate with EcR and USP identified multiple Mediator subunits, including CDK8 and CycC. Consistently, CDK8-CycC interacts with EcR-USP in vivo; in particular, CDK8 and Med14 can directly interact with the AF1 domain of EcR. These results suggest that CDK8-CycC may serve as transcriptional cofactors for EcR-dependent transcription. During the larval-pupal transition, the levels of CDK8 protein positively correlate with EcR and USP levels, but inversely correlate with the activity of sterol regulatory element binding protein (SREBP), the master regulator of intracellular lipid homeostasis. Likewise, starvation of early third instar larvae precociously increases the levels of CDK8, EcR and USP, yet down-regulates SREBP activity. Conversely, refeeding the starved larvae strongly reduces CDK8 levels but increases SREBP activity. Importantly, these changes correlate with the timing for the larval-pupal transition. Taken together, these results suggest that CDK8-CycC links nutrient intake to developmental transitions (EcR activity) and fat metabolism (SREBP activity) during the larval-pupal transition (Xie, 2015).
In animals, the amount of juvenile growth is controlled by the coordinated timing of maturation and growth rate, which are strongly influenced by the environmental factors such as nutrient availability. This is particularly evident in arthropods, such as insects, arachnids and crustaceans, which account for over 80% of all described animal species on earth. Characterized by their jointed limbs and exoskeletons, juvenile arthropods have to replace their rigid cuticles periodically by molting. In insects, the larval-larval and larval-pupal transitions are controlled by the interplay between juvenile hormone (JH) and steroid hormone ecdysone. Drosophila has been a powerful system for deciphering the conserved mechanisms that regulate hormone signaling, sugar and lipid homeostasis, and the molecular mechanisms underlying the nutritional regulation of development. In Drosophila, all growth occurs during the larval stage when larvae constantly feed, and as a result their body mass increases approximately 200-fold within 4 d, largely due to de novo lipogenesis. At the end of the third instar, pulses of ecdysone, combined with a low level of JH, trigger the larval-pupal transition and metamorphosis. During this transition, feeding is inhibited, and after pupariation, feeding is impossible, thus the larval-pupal transition marks when energy metabolism is switched from energy storage by lipogenesis in larvae to energy utilization by lipolysis in pupae (Xie, 2015).
The molecular mechanisms of ecdysone-regulated metamorphosis and developmental timing have been studied extensively in Drosophila. Ecdysone binds to the Ecdysone Receptor (EcR), which heterodimerizes with Ultraspiracle (USP), an ortholog of the vertebrate Retinoid X Receptor (RXR). By activating the expression of genes whose products are required for metamorphosis, ecdysone and EcR-USP are essential for the reorganization of flies' body plans before emerging from pupal cases as adults. Despite the tremendous progress in understanding of the physiological and developmental effects of EcR-USP signaling, the molecular mechanism of how the EcR-USP transcription factor interacts with the general transcription machinery of RNA polymerase II (Pol II) and stimulates its target gene expression remains mysterious. EcR is colocalized with Pol II in Bradysia hygida and Chironomus tentans. Although a number of proteins, such as Alien, Bonus, Diabetes and Obesity Regulated (dDOR), dDEK, Hsc70, Hsp90, Rigor mortis (Rig), Smrter (Smr), Taiman, and Trithorax-related (TRR), have been identified as regulators or cofactors of EcR-mediated gene expression, it is unknown how these proteins communicate with the general transcription machinery and whether additional cofactors are involved in EcR-mediated gene expression. In addition, it remains poorly understood how EcR activates transcription correctly after integrating nutritional and developmental cues (Xie, 2015).
The multisubunit Mediator complex serves as a molecular bridge between transcriptional factors and the core transcriptional machinery, and is thought to regulate most (if not all) of Pol II-dependent transcription. Biochemical analyses have identified two major forms of the Mediator complexes: the large and the small Mediator complexes. In addition to a separable 'CDK8 submodule', the large Mediator complex contains all but one (MED26) of the subunits of the small Mediator complex. The CDK8 submodule is composed of MED12, MED13, CDK8, and CycC. CDK8 is the only enzymatic subunit of the Mediator complex, and CDK8 can both activate and repress transcription depending on the transcription factors with which it interacts. Amplification and mutation of genes encoding CDK8, CycC, and other subunits of Mediator complex have been identified in a variety of human cancers, however, the function and regulation of CDK8-CycC in non-disease conditions remain poorly understood. CDK8 and CycC are highly conserved in eukaryotes, thus analysis of the functional regulation of CDK8-CycC in Drosophila is a viable approach to understand their activities (Xie, 2015).
Previous, work has shown that CDK8-CycC negatively regulates the stability of sterol regulatory element-binding proteins (SREBPs) by directly phosphorylating a conserved threonine residue. This study now reports that CDK8-CycC also regulates developmental timing in Drosophila by linking nutrient intake with EcR-activated gene expression. Homozygous cdk8 or cycC mutants resemble EcR mutants in both pupal morphology and retarded developmental transitions. Despite the elevation of both EcR and USP proteins in cdk8 or cycC mutants, genome-wide gene expression profiling analyses reveal systematic down-regulation of EcR-target genes, suggesting the CDK8-CycC defect lies between the receptor complex and transcriptional activation. CDK8-CycC is required for EcR-USP transcription factor binding to EcR target genes. Mass spectrometry analysis for proteins that co-immunoprecipitate with EcR and USP has identified multiple Mediator subunits, including CDK8 and CycC, and yeast two-hybrid assays have revealed that CDK8 and Med14 can directly interact with the EcR-AF1 domain. Furthermore, the dynamic changes of CDK8, EcR, USP, and SREBP correlated with the fundamental roles of SREBP in regulating lipogenesis and EcR-USP in regulating metamorphosis during the larval-pupal transition. Importantly, it was shown that starving the early third instar larvae causes precocious increase of CDK8, EcR and USP proteins, as well as premature inactivation of SREBP; whereas refeeding of the starved larvae reduces CDK8, EcR, and USP proteins, but potently stimulates SREBP activity. These results suggest a dual role of CDK8-CycC, linking nutrient intake to de novo lipogenesis (by inhibiting SREBP) and developmental signaling (by regulating EcR-dependent transcription) during the larval-pupal transition (Xie, 2015).
Through EcR-USP, ecdysone plays pivotal roles in controlling developmental timing in Drosophila. This study shows that cdk8 or cycC mutants resemble EcR-B1 mutants and CDK8-CycC is required for proper activation of EcR-target genes. Molecular and biochemical analyses suggest that CDK8-CycC and the Mediator complexes are directly involved in EcR-dependent gene activation. In addition, the protein levels of CDK8 and CycC are up-regulated at the onset of the wandering stage, closely correlated with the activation of EcR-USP and down-regulation of SREBP-dependent lipogenesis during the larval-pupal transition. Remarkably, starvation of the feeding larvae leads to premature up-regulation of CDK8 and EcR-USP, and precocious down-regulation of SREBP, while refeeding of the starved larvae results in opposite effects on the CDK8-SREBP/EcR network. Thus, it is proposed that CDK8-CycC serves as a key mediator linking food consumption and nutrient intake to EcR-dependent developmental timing and SREBP-dependent lipogenesis during the larval-pupal transition (Xie, 2015).
The Mediator complex is composed of up to 30 different subunits, and biochemical analyses of the Mediator have identified the small Mediator complex and the large Mediator complex, with the CDK8 submodule being the major difference between the two complexes. Several reports link EcR and certain subunits of the Mediator complex. For example, Med12 and Med24 were shown to be required for ecdysone-triggered apoptosis in Drosophila salivary glands. It was recently reported that ecdysone and multiple Mediator subunits could regulate cell-cycle exit in neuronal stem cells by changing energy metabolism in Drosophila, and specifically, EcR was shown to co-immunoprecipitate with Med27. However, exactly how Mediator complexes are involved in regulating EcR-dependent transcription remains unknown. The current data suggest that CDK8 and CycC are required for EcR-activated gene expression. Loss of either CDK8 or CycC reduced USP binding to EcR target promoters, diminished EcR target gene expression, and delayed developmental transition, which are reminiscent of EcR-B1 mutants. Importantly, mass spectrometry analysis for proteins that co-immunoprecipitate with EcR or USP has identified multiple Mediator subunits, including all four subunits of the CDK8 submodule (Xie, 2015).
Taken together, previous works and the present work highlight a critical role of the Mediator complexes including CDK8-CycC in regulating EcR-dependent transcription.
How does CDK8-CycC regulate EcR-activated gene expression? Biochemical analyses show that CDK8 can interact with EcR and USP in vivo and that CDK8 can directly interact with EcR-AF1. These observations, together with the current understanding of how nuclear receptors and Mediator coordinately regulate transcription, suggest that CDK8-CycC may positively and directly regulate EcR-dependent transcription. Yeast two-hybrid analysis indicates that the recruitment of CDK8-CycC to EcR-USP can occur via interactions between CDK8 and the AF1 domain of EcR. Interestingly, this assay also revealed a direct interaction between EcR-AF1 and a fragment of Med14 that contains the LXXLL motif. In future work, it will be interesting to determine whether CDK8 and Med14 compete with each other in binding with the EcR-AF1, whether they interact with EcR-AF1 sequentially in activating EcR-dependent transcription, and how the Mediator complexes coordinate with other known EcR cofactors in regulating EcR-dependent gene expression (Xie, 2015).
In cdk8 or cycC mutants, the binding of USP to the promoters of the EcR target genes is significantly compromised, even though nuclear protein levels of both EcR and USP are increased. It is unclear how CDK8-CycC positively regulates EcR-USP binding to EcREs near promoters. CDK8 can directly phosphorylate a number of transcription factors, such as Notch intracellular domain, E2F1, SMADs, SREBP, STAT1, and p53. Interestingly, the endogenous EcR and USP are phosphorylated at multiple serine residues, and treatment with 20E enhances the phosphorylation of USP. Protein kinase C has also been proposed to phosphorylate USP. It will be interesting to determine whether CDK8 can also directly phosphorylate either EcR or USP, thereby potentiating expression of EcR target genes and integrating signals from multiple signaling pathways (Xie, 2015).
Although a direct role for CDK8-CycC to regulate EcR-USP activated gene expression is favored, it was not possible to exclude the potential contribution of impaired biosynthesis of 20E to the developmental defects in cdk8 or cycC mutants. For example, the expression of genes involved in synthesis of 20E, such as sad and spok, is significantly reduced in cdk8 or cycC mutant larvae. Indeed, the ecdysteroid titer is significantly lower in cdk8 mutants than control from the early L3 to the WPP stages, and feeding the cdk8 mutant larvae with 20E can partially reduce the retardation in pupariation. Nevertheless, impaired ecdysone biosynthesis alone cannot explain developmental defects that were characterized in this report for the following reasons. First, feeding cdk8 or cycC mutants with 20E cannot rescue the defects in pupal morphology, developmental delay, and the onset of pupariation. Second, the expression of EcRE-lacZ reporter in cdk8 or cycC mutant salivary glands cannot be as effectively stimulated by 20E treatment as in control. Third, knocking down of either cdk8 or cycC in PG did not lead to obvious defects in developmental timing. Therefore, the most likely scenario is that the cdk8 or cycC mutants are impaired not only in 20E biosynthesis in the PG, but also in EcR-activated gene expression in peripheral tissues. Defects in either ecdysone biosynthesis or EcR transcriptional activity will generate the same outcome: diminished expression of the EcR target genes, thereby delayed onset of pupariation (Xie, 2015).
How CDK8-CycC regulates biosynthesis of ecdysone in PG remains unknown. Several signaling pathways have been proposed to regulate ecdysone biosynthesis in Drosophila PG, including PTTH and Drosophila insulin-like peptides (dILPs)-activated receptor tyrosine kinase pathway and Activins/TGFβ signaling pathway. Interestingly, CDK8 has been reported to regulate the transcriptional activity of SMADs, transcription factors downstream of the TGFβ signaling pathway, in both Drosophila and mammalian cells. Thus, it is conceivable that the effect of cdk8 or cycC mutation on ecdysone biosynthesis may due to dysregulated TGFβ signaling in the PG (Xie, 2015).
An effort to explore the potential role of food consumption and nutrient intake on CDK8-CycC has resulted an unexpected observation that the protein level of CDK8 is strongly influenced by starvation and refeeding: starvation potently increased CDK8 level, while refeeding has opposite effect, and both occur post-transcriptionally. The importance of this observation is highlighted in two aspects. First, considering the generally repressive role of CDK8 on Pol II-dependent gene expression, up-regulation of CDK8 may provide an efficient way to quickly tune down most of the Pol II-dependent transcription in response to starvation; while down-regulation of CDK8 in response to refeeding may allow many genes to express when nutrients are abundant. Second, it will be necessary to test whether the effects of nutrient intake on CDK8-CycC is conserved in mammals. If so, considering that both CDK8 and CycC are dysregulated in a variety of human cancers, the effects of nutrient intake on CDK8 may have important implications in not only understanding of the effects of nutrients on tumorigenesis, but also providing nutritional guidance for patients with cancer (Xie, 2015).
Major dietary components including carbohydrates, lipids, and proteins, can strongly influence the developmental timing in Drosophila. Excessive dietary carbohydrates repress growth and potently retard the onset of pupariation. One elegant model proposed to explain how high sugar diet delays developmental timing is that high sugar diet reduces the activity of the Target of Rapamycin (TOR) in the PG, thereby reducing the secretion of ecdysone and delaying the developmental transition. Previously, it was reported that insulin signaling could down-regulate CDK8-CycC, and that ectopic expression of CycC could antagonize the effect of insulin stimulation in mammalian cells, as well as the effect of refeeding on the expression of dFAS in Drosophila (Zhao, 2012). Although the mRNA levels of TOR and insulin receptor (InR) are not significantly affected in cdk8 or cycC mutants, it is necessary to further study whether and how different dietary components may regulate CDK8-CycC in the future (Xie, 2015).
Developmental genetic analyses of the cdk8 and cycC mutants have revealed major defects in fat metabolism and developmental timing. De novo lipogenesis, which is stimulated by insulin signaling, contributes significantly to the rapid increase of body mass during the constant feeding larval stage. This process is terminated by pulses of ecdysone that trigger the wandering behavior at the end of the L3 stage, followed by the onset of the pupariation. Insulin and ecdysone signaling are known to antagonize each other, and together determine body size of Drosophila. The genetic interaction is established, but the detailed molecular mechanisms are not. The SREBP family of transcription factors controls the expression of lipogenic enzymes in metazoans and the expression of cholesterogenic enzymes in vertebrates. Previous work shows that CDK8 directly phosphorylates the nuclear SREBP proteins on a conserved threonine residue and promotes the degradation of nuclear SREBP proteins. Consistent with the lipogenic role of SREBP and the inhibitory role of insulin to CDK8-CycC, the transcriptional activity of SREBP is high while the levels of CDK8-CycC and EcR-USP are low prior to the onset of wandering stage. Subsequently during the wandering and non-mobile, non-feeding pupal stage, the transcriptional activity of SREBP is dramatically reduced, accompanied by the significant accumulation of CDK8-CycC and EcR-USP (Xie, 2015).
The causal relationship of these phenomena was further tested by starvation and refeeding experiments. On the one hand, it was observed that the levels of CDK8, EcR and USP are potently induced by starvation, while the mature SREBP level and the transcriptional activity of SREBP are reduced by starvation. Starvation of larvae prior to the two nutritional checkpoints in early L3, known as minimum viable weight and critical weight, which are reached almost simultaneously in Drosophila, will lead to larval lethality; while starvation after larvae reach the critical weight will lead to early onset of pupariation and formation of small pupae. Thus, this nutritional checkpoint ensures the larvae have accumulated sufficient growth before metamorphosis initiation. If the status with high CDK8, EcR, and USP is regarded as an older or later stage, these results indicate that starvation shifts the regulatory network precociously (see Model for the CDK8-SREBP/EcR regulatory network). On the other hand, the current analyses of refed larvae show that refeeding potently reduced the levels of CDK8, EcR and USP. If the status with low CDK8, EcR, and USP is considered as a younger or earlier stage, these results indicate that refeeding delays the activation of this network, which is consistent with the model and delayed pupariation as observed. Taken together, these results based on starved and refed larvae suggest that CDK8-CycC is a key regulatory node linking nutritional cues with de novo lipogenesis and developmental timing (Xie, 2015).
The larval-pupal transition is complex and dynamic. Although the expression of SREBP target genes fit well with the predicted effects of starvation and refeeding, the expression of EcR targets during the stage that was analyzed in this study does not reflect the changes in the protein levels of EcR and USP. It is reasonable to consider that CDK8-CycC and EcR-USP are necessary, but not sufficient, for the activation of EcR target genes. One possibility is that there is a delay on synthesis of 20E or other cofactors that are required for EcR-activated gene expression in response to starvation. Indeed, the 20E levels were measured during the first 16 hr of starvation, and no significant difference was observed between fed and starved larvae. It will be necessary to further analyze the effect of starvation on 20E synthesis at later time points in the future (Xie, 2015).
Taken together, a model is proposed whereby CDK8-CycC functions as a regulatory node that coordinates de novo lipogenesis during larval stage and EcR-dependent pupariation in response to nutritional cues. It is likely that pulses of 20E synthesized in the PG, and subsequent behavioral change from feeding to wandering, ultimately trigger the transition from SREBP-dependent lipogenesis to EcR-dependent pupariation. The opposite effects of CDK8-CycC on SREBP- and EcR-dependent gene expression suggest that the role of CDK8 on transcription is context-dependent (Xie, 2015).
In conclusion, this study illustrates how CDK8-CycC regulates EcR-USP-dependent gene expression, and the results suggest that CDK8-CycC may function as a regulatory node linking fat metabolism and developmental timing with nutritional cues during Drosophila development (Xie, 2015).
Body shapes are much more variable than body plans. One way to alter body shapes independently of body plans would be to mechanically deform bodies. To what extent body shapes are regulated physically, or molecules involved in physical control of morphogenesis, remain elusive. During fly metamorphosis, the cuticle (exoskeleton) covering the larval body contracts longitudinally and expands laterally to become the ellipsoidal pupal case (puparium). This study shows that Drosophila melanogaster Obstructor-E (Obst-E) is a protein constituent of the larval cuticle that confers the oriented contractility/expandability. In the absence of obst-E function, the larval cuticle fails to undergo metamorphic shape change and finally becomes a twiggy puparium. Results are presented indicating that Obst-E regulates the arrangement of chitin, a long-chain polysaccharide and a central component of the insect cuticle, and directs the formation of supracellular ridges on the larval cuticle. It was further shown that Obst-E is locally required for the oriented shape change of the cuticle during metamorphosis, which is associated with changes in the morphology of those ridges. Thus, Obst-E dramatically affects the body shape in a direct, physical manner by controlling the mechanical property of the exoskeleton (Tajiri, 2017).
The central mechanisms coordinating growth and sexual maturation are well conserved across invertebrates and vertebrates. Although mutations in the gene encoding makorin RING finger protein 3 (mkrn3) are associated with central precocious puberty in humans, a causal relationship has not been elucidated. This study examined the role of the
Drosophila ortholog of mammalian makorin genes, in the regulation of developmental timing. Loss of MKRN1 in mkrn1exS prolonged the 3rd instar stage and delayed the onset of pupariation, resulting in bigger size pupae. MKRN1 was expressed in the prothoracic gland, where the steroid hormone ecdysone is produced. Furthermore, mkrn1exS larvae exhibited reduced mRNA levels of phantom, which encodes ecdysone-synthesizing enzyme and E74, which is a downstream target of ecdysone. Collectively, these results indicate that MKRN1 fine-tunes developmental timing and sexual maturation by affecting ecdysone synthesis in Drosophila. Moreover, this study supports the notion that malfunction of makorin gene family member, mkrn3 dysregulates the timing of puberty in mammals (Tran, 2018).
During morphogenesis, cells exhibit various behaviours, such as migration and constriction, which need to be coordinated. How this is achieved remains elusive. During morphogenesis of the Drosophila adult abdominal epidermis, larval epithelial cells (LECs) migrate directedly before constricting apically and undergoing apoptosis. This study investigates the mechanisms underlying the transition from migration to constriction. LECs possess a pulsatile apical actomyosin network, and a change in network polarity correlates with behavioural change. Exploring the properties of the contractile network, it was found that cell contractility, as determined by myosin activity, has an impact on the behaviour of the network, as well as on cytoskeletal architecture and cell behaviour. Pulsed contractions occur only in cells with intermediate levels of contractility. Furthermore, increasing levels of the small Rho GTPase Rho1 disrupts pulsing, leading to cells that cycle between two states, characterised by a junctional cortical and an apicomedial actin network. These results highlight that behavioural change relies on tightly controlled cellular contractility. Moreover, constriction can occur without pulsing, raising questions why constricting cells pulse in some contexts but not in others (Companys, 2020).
In holometabolous insects, the growth period is terminated through a cascade of peptide and steroid hormones that end larval feeding behavior and trigger metamorphosis, a nonfeeding stage during which the larval body plan is remodeled to produce an adult. This irreversible decision, termed the critical weight (CW) checkpoint, ensures that larvae have acquired sufficient nutrients to complete and survive development to adulthood. How insects assess body size via the CW checkpoint is still poorly understood on the molecular level. This study shows that the Drosophila transcription factor Snail plays a key role in this process. Before and during the CW checkpoint, snail is highly expressed in the larval prothoracic gland (PG), an endocrine tissue undergoing endoreplication and primarily dedicated to the production of the steroid hormone ecdysone. Two Snail peaks were observed in the PG, one before and one after the molt from the second to the third instar. Remarkably, these Snail peaks coincide with two peaks of PG cells entering S phase and a slowing of DNA synthesis between the peaks. Interestingly, the second Snail peak occurs at the exit of the CW checkpoint. Snail levels then decline continuously, and endoreplication becomes nonsynchronized in the PG after the CW checkpoint. This suggests that the synchronization of PG cells into S phase via Snail represents the mechanistic link used to terminate the CW checkpoint. Indeed, PG-specific loss of snail function prior to the CW checkpoint causes larval arrest due to a cessation of endoreplication in PG cells, whereas impairing snail after the CW checkpoint no longer affected endoreplication and further development. During the CW window, starvation or loss of TOR signaling disrupted the formation of Snail peaks and endocycle synchronization, whereas later starvation had no effect on snail expression. Taken together, these data demonstrate that insects use the TOR pathway to assess nutrient status during larval development to regulate Snail in ecdysone-producing cells as an effector protein to coordinate endoreplication and CW attainment (Zeng, 2020).
Metabolism must be coupled with developmental transition to fulfill the energy requirements during an organism's life cycle. In most animals, steroid hormones are crucial regulators of life-stage transitions until adulthood. In the fruit fly Drosophila, ecdysteroid titers drive developmental transitions, such as molting and metamorphosis. Although the timings of molting and larval-pupal transition are dependent on environmental and internal conditions, metamorphosis is a temporally controlled life transition event that solely relies on energetic macromolecules accumulated during the larval period. The ecdysteroid signaling cascade and the energetics of metamorphosis have been determined. However, the molecular mechanisms that regulate metabolic homeostasis during metamorphosis remain largely unknown. This study shows that the programmed regulation of carbohydrate metabolism by steroid hormones directs the prepupal-pupal transition in Drosophila. Pupation is associated with a transient increase in glucose oxidation. Mechanistically, after pupariation, ecdysteroid signaling and the competence factor Ftz-F1 regulates the systematic degradation of circulating trehalose via the transient induction of trehalose transporters and trehalase (Treh) in a timely manner. Trehalose metabolism is crucial for energy homeostasis at the prepupal-pupal transition. Moreover, trehalose catabolism acts upstream of ecdysteroid biosynthesis and signaling. These findings lead to the hypothesis that trehalose breakdown not only defines energy costs by providing a carbon source but also facilitates pupation by inducing water loss in the puparium. This work sheds light on the ways in which a life-stage transition is driven by the preprogrammed coordination between steroid hormones and catabolism of stored nutrients (Nishimura, 2020).
The daily rhythm of adult emergence of holometabolous insects is one of the first circadian rhythms to be studied. In these insects, the circadian clock imposes a daily pattern of emergence by allowing or stimulating eclosion during certain windows of time and inhibiting emergence during others, a process that has been described as "gating." Although the circadian rhythm of insect emergence provided many of the key concepts of chronobiology, little progress has been made in understanding the bases of the gating process itself, although the term "gating" suggests that it is separate from the developmental process of metamorphosis. This study followed the progression through the final stages of Drosophila adult development with single-animal resolution and showed that the circadian clock imposes a daily rhythmicity to the pattern of emergence by controlling when the insect initiates the final steps of metamorphosis itself. Circadian rhythmicity of emergence depends on the coupling between the central clock located in the brain and a peripheral clock located in the prothoracic gland (PG), an endocrine gland whose only known function is the production of the molting hormone, ecdysone. This study shows that the clock exerts its action by regulating not the levels of ecdysone but that of its actions mediated by the ecdysone receptor. These findings may also provide insights for understanding the mechanisms by which the daily rhythms of glucocorticoids are produced in mammals, which result from the coupling between the central clock in the suprachiasmatic nucleus and a peripheral clock located in the suprarenal gland (Mark, 2021).
Pulses of the steroid hormone ecdysone act through transcriptional cascades to direct the major developmental transitions during the Drosophila life cycle. These include the prepupal ecdysone pulse, which occurs 10 h after pupariation and triggers the onset of adult morphogenesis and larval tissue destruction. E93 encodes a transcription factor that is specifically induced by the prepupal pulse of ecdysone, supporting a model proposed by earlier work that it specifies the onset of adult development. Although a number of studies have addressed these functions for E93, little is known about its roles in the salivary gland where the E93 locus was originally identified. This study shows that E93 is required for development through late pupal stages, with mutants displaying defects in adult differentiation and no detectable effect on the destruction of salivary glands. RNA-seq analysis demonstrates that E93 regulates genes involved in development and morphogenesis in the salivary glands, but has little effect on cell death gene expression. It was also shown that E93 is required to direct the proper timing of ecdysone-regulated gene expression in salivary glands, and that it suppresses earlier transcriptional programs that occur during larval and prepupal stages. These studies support the model that the stage-specific induction of E93 in late prepupae provides a critical signal that defines the end of larval development and the onset of adult differentiation (Lam, 2021).
The genome of insects with complete metamorphosis contains the instructions for making three distinct body forms, that of the larva, of the pupa, and of the adult. However, the molecular mechanisms by which each gene set is called forth and stably expressed are poorly understood. A half century ago, it was proposed that there was a set of three master genes that inhibited each other's expression and enabled the expression of genes for each respective stage. This study shows that the transcription factor chinmo is essential for maintaining the larval stage in Drosophila, and with two other regulatory genes, broad and E93, makes up the trinity of mutually repressive master genes that underlie insect metamorphosis (Truman, 2022).
For in vivo functional analysis of a protein of interest (POI), multiple transgenic strains with a POI that harbors different tags are needed but generation of these strains is still labor-intensive work. To overcome this, a versatile Drosophila toolkit was developed with a genetically encoded single-chain variable fragment for the HA epitope tag: 'HA Frankenbody'. This system allows various analyses of HA-tagged POI in live tissues by simply crossing an HA Frankenbody fly with an HA-tagged POI fly. Strikingly, the GFP-mCherry tandem fluorescent-tagged HA Frankenbody revealed a block in autophagic flux and an accumulation of enlarged autolysosomes in the last instar larval and prepupal fat body. Mechanistically, lysosomal activity was downregulated at this stage, and endocytosis, but not autophagy, was indispensable for the swelling of lysosomes. Furthermore, forced activation of lysosomes by fat body-targeted overexpression of Mitf, the single MiTF/TFE family gene in Drosophila, suppressed the lysosomal swelling and resulted in pupal lethality. Collectively, it is proposed that downregulated lysosomal function in the fat body plays a role in the metamorphosis of Drosophila (Murakawa, 2022).
The digestion of food and absorption of nutrients occurs in the gut. The nutritional value of food and its nutrients is detected by enteroendocrine cells, and peptide hormones produced by the enteroendocrine cells are thought to be involved in metabolic homeostasis, but the specific mechanisms are still elusive. The enteroendocrine cells are scattered over the entire gastrointestinal tract and can be classified according to the hormones they produce. This study followed the changes in combinatorial expression of regulatory peptides in the enteroendocrine cells during metamorphosis from the larva to the adult fruit fly, and re-confirmed the diverse composition of enteroendocrine cell populations. Drosophila enteroendocrine cells appear to differentially regulate peptide expression spatially and temporally depending on midgut region and developmental stage. In the late pupa, Notch activity is known to determine which peptides are expressed in mature enteroendocrine cells of the posterior midgut; it was found that the loss of Notch activity in the anterior midgut results in classes of enteroendocrine cells distinct from the posterior midgut. These results suggest that enteroendocrine cells that populate the fly midgut can differentiate into distinct subtypes that express different combinations of peptides, which likely leads to functional variety depending on specific needs (Jang, 2021).
Insect metamorphosis is triggered by the production, secretion and degradation of 20-hydroxyecdysone (ecdysone). In addition to its role in developmental regulation, increasing evidence suggests that ecdysone is involved in innate immunity processes, such as phagocytosis and the induction of antimicrobial peptide (AMP) production. AMP regulation includes systemic responses as well as local responses at surface epithelia that contact with the external environment. At pupariation, Drosophila melanogaster increases dramatically the expression of three AMP genes, drosomycin (drs), drosomycin-like 2 (drsl2) and drosomycin-like 5 (drsl5). The systemic action of drs at pupariation is dependent on ecdysone signalling in the fat body and operates via the ecdysone downstream target, Broad. In parallel, ecdysone also regulates local responses, specifically through the activation of drsl2 expression in the gut. Finally, the relevance of this ecdysone dependent AMP expression for the control of bacterial load was confirmed by showing that flies lacking drs expression in the fat body have higher bacterial persistence over metamorphosis. In contrast, local responses may be redundant with the systemic effect of drs since reduction of ecdysone signalling or of drsl2 expression has no measurable negative effect on bacterial load control in the pupa. Together, these data emphasize the importance of the association between ecdysone signalling and immunity using in vivo studies and establish a new role for ecdysone at pupariation, which impacts developmental success by regulating the immune system in a stage-dependent manner. It is speculated that this co-option of immune effectors by the hormonal system may constitute an anticipatory mechanism to control bacterial numbers in the pupa, at the core of metamorphosis evolution (Nunes, 2021).
Ecdysone, an essential insect steroid hormone, promotes larval metamorphosis by coordinating growth and maturation. In Drosophila melanogaster, prothoracicotropic hormone (PTTH)-releasing neurons are considered to be the primary promoting factor in ecdysone biosynthesis. Recently, studies have reported that the regulatory mechanisms of PTTH release in Drosophila larvae are controlled by different neuropeptides, including allatostatin A and corazonin. However, it remains unclear whether neurotransmitters provide input to PTTH neurons and control the metamorphosis in Drosophila larvae. This study reports that the neurotransmitters acetylcholine (ACh) affect larval development by modulating the activity of PTTH neurons. By downregulating the expression of different subunits of nicotinic ACh receptors in PTTH neurons, pupal volume was significantly increased, whereas pupariation timing was relatively unchanged. It was also identified that PTTH neurons were excited by ACh application ex vivo in a dose-dependent manner via ionotropic nicotinic ACh receptors. Moreover, in Ca(2+) imaging experiments, relatively low doses of OA caused increased Ca(2+) levels in PTTH neurons, whereas higher doses led to decreased Ca(2+) levels. It was also demonstrated that a low dose of OA was conveyed through OA *betal-type receptors. Additionally, electrophysiological experiments revealed that PTTH neurons produced spontaneous activity in vivo, which provides the possibility of the bidirectional regulation, coming from neurons upstream of PTTH cells in Drosophila larvae. In summary, these findings indicate that several different neurotransmitters are involved in the regulation of larval metamorphosis by altering the activity of PTTH neurons in Drosophila (Hao, 2021).
During tissue closures, such as embryonic dorsal closure in Drosophila melanogaster, a proximate extra-embryonic layer, amnioserosa, generates forces that drive migration of the flanking lateral embryonic epidermis, thereby zip-shutting the embryo. Arguably, this paradigm of tissue closure is also recapitulated in mammalian wound healing wherein proximate fibroblasts transform into contractile myofibroblasts, develop cell junctions, and form a tissue layer de novo: contraction of the latter then aids in wound closure. Given this parallelism between disparate exemplars, a general principle of tissue closure is posited via proximate cell layer-generated forces. This hypothesis was tested in pupal thorax closure wherein 2 halves of the presumptive adult thorax of Drosophila, the contralateral heminotal epithelia, migrate over an underlying larval epidermal cell layer. The proximate larval epidermal cell layer promotes thorax closure by its active contraction, orchestrated by its elaborate actomyosin network-driven epithelial cell dynamics, cell delamination, and death-the latter being prefigured by the activation of caspases. Larval epidermal cell dynamics generate contraction forces, which when relayed to the flanking heminota-via their mutual integrin-based adhesions-mediate thorax closure. Compromising any of these contraction force-generating mechanisms in the larval epidermal cell layer slows down heminotal migration, while loss of its relay to the flanking heminota abrogates the thorax closure altogether. Mathematical modeling further reconciles the biophysical underpinning of this emergent mechanism of thorax closure. Revealing mechanism of thorax closure apart, these findings show conservation of an essential principle of a proximate cell layer-driven tissue closure (Athilingam, 2022).
For centuries, it has been understood that the final size of adult holometabolous insects is determined by the end of the larval stage, and that once they transform to adults, holometabolous insects do not grow. Despite this, no previous study has directly tested these 'old truths' across holometabolous insects. This study demonstrates that final adult size is set at the end of the last larval stage in species representing each of the four orders of holometabolous insects: the fruit fly Drosophila melanogaster (Diptera), the tobacco hornworm Manduca sexta (Lepidoptera), the dung beetle Onthophagus taurus (Coleoptera), and the Florida carpenter ant Camponotus floridanus (Hymenoptera). Furthermore, in both D. melanogaster and C. floridanus, this study showed that the size of adult individuals fluctuates but does not significantly change. Therefore, this study finally confirms these two basic assumptions in the biology of insects, which have for centuries served as the foundation for studies of insect growth, size, and allometry (Hanna, 2022).
Pruning that selectively eliminates unnecessary or incorrect neurites is required for proper wiring of the mature nervous system. During Drosophila metamorphosis, dendritic arbourization sensory neurons (ddaCs) and mushroom body (MB) γ neurons can selectively prune their larval dendrites and/or axons in response to the steroid hormone ecdysone. An ecdysone-induced transcriptional cascade plays a key role in initiating neuronal pruning. However, how downstream components of ecdysone signalling are induced remains not entirely understood. This study identified that Scm, a component of Polycomb group (PcG) complexes, is required for dendrite pruning of ddaC neurons. Two PcG complexes, PRC1 and PRC2, are important for dendrite pruning. Interestingly, depletion of PRC1 strongly enhances ectopic expression of Abdominal B (Abd-B) and Sex combs reduced, whereas loss of PRC2 causes mild upregulation of Ultrabithorax and Abdominal A in ddaC neurons. Among these Hox genes, overexpression of Abd-B causes the most severe pruning defects, suggesting its dominant effect. Knockdown of the core PRC1 component Polyhomeotic (Ph) or Abd-B overexpression selectively downregulates Mical expression, thereby inhibiting ecdysone signalling. Finally, Ph is also required for axon pruning and Abd-B silencing in MB γ neurons, indicating a conserved function of PRC1 in two types of pruning. This study demonstrates important roles of PcG and Hox genes in regulating ecdysone signalling and neuronal pruning in Drosophila. Moreover, our findings suggest a non-canonical and PRC2-independent role of PRC1 in Hox gene silencing during neuronal pruning (Bu, 2023).
Drosophila blood cells called hemocytes form an efficient barrier against infections and tissue damage. During metamorphosis, hemocytes undergo tremendous changes in their shape and behavior, preparing them for tissue clearance. Yet, the diversity and functional plasticity of pupal blood cells have not been explored. This study combine single-cell transcriptomics and high-resolution microscopy to dissect the heterogeneity and plasticity of pupal hemocytes. We identified undifferentiated and specified hemocytes with different molecular signatures associated with distinct functions such as antimicrobial, antifungal immune defense, cell adhesion or secretion. Strikingly, a highly migratory and immune-responsive pupal cell population was identified expressing typical markers of the posterior signaling center (PSC), which is known to be an important niche in the larval lymph gland. PSC-like cells become restricted to the abdominal segments and are morphologically very distinct from typical Hemolectin (Hml)-positive plasmatocytes. G-TRACE lineage experiments further suggest that PSC-like cells can transdifferentiate to lamellocytes triggered by parasitoid wasp infestation. In summary, this study presents the first molecular description of pupal Drosophila blood cells, providing insights into blood cell functional diversification and plasticity during pupal metamorphosis (Moore, 2023).
Drosophila blood cells called hemocytes form an efficient barrier against infections and tissue damage. During metamorphosis, hemocytes undergo tremendous changes in their shape and behavior, preparing them for tissue clearance. Yet, the diversity and functional plasticity of pupal blood cells have not been explored. This study combine single-cell transcriptomics and high-resolution microscopy to dissect the heterogeneity and plasticity of pupal hemocytes. This study identified undifferentiated and specified hemocytes with different molecular signatures associated with distinct functions such as antimicrobial, antifungal immune defense, cell adhesion or secretion. Strikingly, a highly migratory and immune-responsive pupal cell population was identified expressing typical markers of the posterior signaling center (PSC), which is known to be an important niche in the larval lymph gland. PSC-like cells become restricted to the abdominal segments and are morphologically very distinct from typical Hemolectin (Hml)-positive plasmatocytes. G-TRACE lineage experiments further suggest that PSC-like cells can transdifferentiate to lamellocytes triggered by parasitoid wasp infestation. In summary, this study presents the first molecular description of pupal Drosophila blood cells, providing insights into blood cell functional diversification and plasticity during pupal metamorphosis (Hirschhauser, 2023).
A small number of signaling molecules, used reiteratively, control differentiation programs, but the mechanisms that adapt developmental timing to environmental cues are less understood. This study reports that a macrophage inr/dtor/pvf2
An understanding of the molecular basis of the endocrine control of insect metamorphosis has been hampered by the profound differences in the responses of the Lepidoptera and the Diptera to juvenile hormone (JH). In the presence of JH,
there is no change in form; in the absence of JH, ecdysone causes the switching in gene expression necessary for metamorphosis, first to the pupa, then to the adult.
JH therefore prevents this switching action of ecdysone and thus maintains the 'status quo' during a molt. In the Coleoptera and in Lepidoptera
such as the tobacco hornworm, Manduca sexta, where the epidermis sequentially makes several larval cuticles, the pupal cuticle and finally the adult cuticle, JH prevents each of the metamorphic transitions. By contrast, in Drosophila and the other higher flies, the pupal epidermis, except
for the abdomen, is derived from imaginal discs, and exogenous JH does not prevent the larval-pupal transformation, even when given throughout larval life. Nor does JH have any effect on the subsequent external adult differentiation of the head and
thorax, although JH disrupts metamorphosis of the nervous and muscular systems when given during the prepupal period. However, JH application during the final larval instar or during the prepupal period prevents the normal adult differentiation of the abdomen, whose cells arise from proliferation of the histoblasts during the prepupal period (Zhou, 2002 and references therein).
In both Manduca and Drosophila, the broad (br) gene is expressed in the epidermis during the formation of the pupa, but not during adult differentiation. Misexpression of Br-Z1 during either a larval or an adult molt of Drosophila suppresses stage-specific cuticle genes and activates pupal cuticle genes, showing that br is a major specifier of the pupal stage.
Treatment with a JH mimic at the onset of the adult molt causes br re-expression and the formation of a second pupal cuticle in Manduca, but only in
the abdomen of Drosophila. Expression of the Br isoforms during adult development of Drosophila suppresses bristle and hair formation when
induced early or redirects cuticle production toward the pupal program when induced late. Expression of Br-Z1 at both of these times mimics the
effect of JH application but, unlike JH, it causes production of a new pupal cuticle on the head and thorax as well as on the abdomen. Consequently,
the 'status quo' action of JH on the pupal-adult transformation is mediated by the JH-induced re-expression of Br (Zhou, 2002).
Br has long been known to be required for the onset of metamorphosis of Drosophila because the nonpupariating (npr) alleles lack all Br proteins and remain as final instar larvae. In both Drosophila and Manduca, Br transcripts and proteins are expressed prominently during the larval-pupal transformation with different isoforms showing different temporal and tissue specificities through this period and causing either activation or suppression of specific genes. For example, in the Drosophila salivary gland, the induction of Sgs-4 and L71 and the suppression of Pig-1 during the mid and late third instar require the Z1 isoform, while the later suppression of Sgs-4 at puparium formation is due to the downregulation of another transcription factor Forkhead (Fkh) by the Z3 isoform. By contrast, the Z3 isoform activates the expression of Fbp1 in larval fat bodies during the second half of the third instar, while Z2 may play a role in repressing its premature expression. Br proteins also may play a role in the regulation of chromatin structure, since they are found in over 300 sites on the salivary gland chromosomes including sites in the interband regions and in the heterochromatin (Zhou, 2002).
Br-Z1 is the predominant isoform during the time of pupal cuticle formation in Drosophila. Whenever Br-Z1 is expressed during an ecdysone-induced molt, it can direct the epidermis into a program of pupal cuticle production. For example, the molt to the third larval instar in Drosophila begins with the rise of the ecdysteroid that peaks about 12 hours after ecdysis to the second instar. Shortly thereafter, mRNAs for larval cuticular proteins are upregulated. Expression of Br-Z1 during this time suppresses the larval cuticle gene Lcp65A-b and prematurely activates the pupal cuticle gene Edg78E. The ability of Br to be a pupal specifier is also evident during an adult molt. This molt begins about 24 hour APF with the rise of the ecdysteroid titer, and adult procuticle deposition begins about 53 hours APF during the decline of the ecdysteroid titer. Br-Z1 is most effective in activating pupal cuticle genes and suppressing an adult cuticle gene when expressed just before the normal onset of adult procuticle gene expression. This temporal restriction suggests that although Br selects which cuticle genes will be expressed, it can only do so within the confines of an ecdysone-induced program that determines the timing of cuticle gene expression at every molt. Therefore, in either a larval or an adult molt, the expression of Br-Z1 is sufficient to redirect that molt towards the pupal program (Zhou, 2002).
Adult differentiation of the epidermis can be divided into two developmental phases: cellular morphogenesis followed by cuticle deposition. Morphogenesis of the epidermis begins with the formation and outgrowth of the bristles (macrochaetes, microchaetes) between 30 and 45 hours APF, first in the head and thorax, then in the abdomen. Trichomes (hairs) are then formed by most of the general epidermal cells, beginning on the wing at 33 hours APF and on the abdomen about 48 hours APF. The general epidermis deposits three cuticular layers: cuticulin, epicuticle and procuticle. The bristle and hair shafts lack the procuticle layer. Cuticulin formation begins in patches on the wings and legs at 35-36 hours and on the abdomen at 40-45 hours APF, followed by synthesis of a continuous epicuticle once morphogenesis is complete. Adult procuticle synthesis occurs primarily between 53 and 90 hours APF. The expression of the adult cuticle gene Acp65A is restricted to flexible cuticle regions of the abdomen, the wing hinges, leg joints and the ptilinum and begins about 55-60 hours APF (Zhou, 2002).
Br disappears before the onset of adult differentiation in both Manduca and Drosophila. This disappearance is crucial for normal adult development since the misexpression of Br in Drosophila can affect both adult morphogenesis and adult cuticle production. When expressed between 30 and 40 hours APF, Br causes truncation of the bristles with early times affecting the bristles of the head and thorax and slightly later times affecting those of the abdomen. This timing corresponds to the onset of bristle outgrowth in the different regions. Suppression of bristle outgrowth occurs with misexpression of each of the Br isoforms, although the Z1 isoform has the strongest effects because the truncation is seen with expression of only two copies as well as with four copies of Br-Z1. Bristle outgrowth occurs by extension of the longitudinal actin microfilament arrays that surround the microtubular core. These actin filaments are bundled together, then crosslinked to support the elongating bristles, using sequentially the product of the forked gene and fascin. Although an occasional forked bristle is seen after misexpression of Br, the primary effect is truncation similar to that seen after exposure to inhibitors of microfilament elongation, indicating Br may be able to interfere with this process, either directly or indirectly (Zhou, 2002).
Trichome production in the abdominal epidermis is suppressed by Br-Z1 expression between 36 and 39 hours APF. Since nearly every epidermal cell normally produces a trichome, this result shows that early Br expression also suppresses morphogenesis of the general epidermis. In this case, the effective time is about 10-12 hours before abdominal trichome production. By 42 hours APF bristle and trichome morphogenesis is no longer affected by expression of Br. Between this time and 60 hours APF, the effects are primarily on the types of cuticle proteins produced. Br-Z1 is most effective in suppressing adult cuticle gene expression and causing re-induction of pupal cuticle gene expression with the resultant formation of a thin, transparent, pupal-like cuticle by the general epidermis. None of the other isoforms have such a dramatic effect on the external appearance of the cuticle, although Br-Z2 causes re-expression of the two pupal cuticle genes, and Br-Z3 causes re-expression of one pupal cuticle gene and suppression of the adult cuticle gene studied, indicating that they normally play a role in production of pupal cuticle. Cuticle is composed of many proteins, so a predominance of adult cuticle proteins could maintain the cuticular morphology despite the presence of some pupal cuticle proteins or the absence of specific adult cuticle proteins. Further study is required to resolve this issue (Zhou, 2002).
Although bristle morphogenesis is unaffected by expression of Br during the onset of cuticle formation, bristle pigmentation and sclerotization are subsequently inhibited. Whether this suppression is due to the type of epicuticle deposited or to an inhibitory action of Br on the melanization and sclerotization pathways themselves is unclear. In the case of Br-Z1, this effect is most pronounced when expression is either between 43 and 48 hours APF or later during 54-60 hours APF. Although the pupal cuticle genes used in this study all encode proteins found in the pupal exocuticle (the outer procuticle), Br-Z1 probably also directs pupal epicuticle production. If so, the earlier expression of Br-Z1 may be suppressing the deposition of the proenzymes necessary for tanning and melanization that are normally associated with adult cuticle. Such a suppression would not be unexpected, since normal pupal cuticle does not tan or melanize. These proenzymes are often laid down very early in formation of the new cuticle. Br-Z3 or Br-Z4 also suppresses bristle pigmentation but only when expressed late between 52 and 60 hours APF. This effect of later expression of any of these three isoforms is probably due to an interference with the production or deposition of the substrates for these enzymes, which normally appear in the cuticle shortly before the proenzymes are activated. However, an effect on the pigmentation process itself that occurs later cannot be ruled out (Zhou, 2002).
These different effects of Br misexpression depending on its timing indicate that Br and/or the unknown proteins whose expression Br regulates must be present to direct the pupal program. Once they disappear, the cells can revert back to the expression of the adult program. In these experiments, Br transcripts disappear by 6 hours after the heat shock, but the proteins are present until at least 9 hours. Thus, in order to obtain a second pupal cuticle that lacks bristles and trichomes, one must express Br-Z1 during both the initiation of bristle outgrowth and the onset of procuticle formation (Zhou, 2002).
An important finding of these studies is the fact that the presence of Br-Z1 at the time of cuticle formation is sufficient to redirect the program of cuticle gene expression into a pupal mode in cells that have completed their adult morphogenesis. This is most clearly seen after expression of Br at 48 or 52 hours APF. The cells of the general abdominal epidermis make the adult hairs but then deposit procuticle that includes pupal cuticle proteins. Thus, cells already committed to and expressing aspects of adult differentiation are plastic and can be caused to re-express pupal products when given the proper transcription factor. Clearly the suppression of br through the duration of adult development is essential for the normal completion of metamorphosis (Zhou, 2002).
JH has long been known to prevent metamorphosis without interfering with the molting process itself. In both Manduca and Drosophila abdomens, JH causes the formation of a second pupal cuticle only when given before the onset of the adult molt. These studies have revealed that this re-expression of the pupal program in both species is associated with the re-induction and maintenance of Br expression during the molt. This renewed Br expression appears to be sufficient to mediate the 'status quo' action of JH, since Br can both activate pupal genes and suppress adult genes. Thus, during the crucial period of adult commitment, ecdysone in the absence of JH must switch off Br so that the adult-specific program of differentiation can occur (Zhou, 2002).
In Drosophila the JH-sensitive period of the abdomen is during the prepupal period with the highest sensitivity being at the time of pupariation and loss of sensitivity after head eversion at 12 hours APF. During this time the histoblasts are proliferating rapidly. After this JH-sensitive period is over, beginning about 15 hours APF, these imaginal cells spread over the pupal abdomen and replace most of the larval cells by about 28 hours APF. Throughout this period, both types of cells express Br. JH given at pupariation has no apparent effect on the proliferation or spreading of these cells or on their replacement of the larval epidermis. Nor does it interfere with their normal Br expression during this period. Its effect is only to cause renewed and sustained expression of Br in the imaginal cells during the adult molt up through 72 hours APF (Zhou, 2002).
JH at pupariation has no apparent effect on the adult development of the Drosophila head and thoracic structures that are derived from the imaginal discs. This study shows that the refractoriness of the head and thorax to the JH treatment is due to the inability of JH to cause Br re-expression in these regions during the adult molt. Yet appropriate misexpression of Br during adult differentiation results in pupal cuticle formation in both the head and thorax, showing that Br retains its pupal-specifying function in these regions. Hence, the refractoriness to JH of the head and thorax must be due to a lesion in the pathway from the JH receptor to Br re-induction, possibly to the loss of the receptor itself (Zhou, 2002).
In all insects including Drosophila, JH is present during the larval molts, then declines during the last larval instar. In both Manduca and Drosophila epidermis and imaginal discs, Br is not expressed during the larval molt. Pupal commitment of the polymorphic epidermis of Manduca by 20E at the end of the larval feeding period is correlated with the appearance of Br, and both can be prevented by JH. By contrast, in Manduca wing imaginal discs, Br appears earlier in the final larval instar as the discs become competent to metamorphose, and JH cannot prevent this appearance but only delays it. In Drosophila and the higher flies, the pupa is derived from imaginal discs except for the abdominal cuticle that is produced by the persisting larval epidermal cells and the histoblasts. Although the effect of JH on the appearance of Br in Drosophila discs and larval epidermis has not been directly studied, dietary JH throughout larval life delays the onset of metamorphosis but does not prevent pupation, indicating that these tissues can turn on Br despite the presence of JH. Thus, the derivation of the Drosophila pupa from primarily imaginal discs probably accounts for the inability of JH to prevent the larval-pupal transformation, although the lack of effect of JH on the abdominal epidermis in its switch to pupal cuticle production remains unexplained. The mechanism whereby JH prevents the switching-on of Br by ecdysone during a larval molt and also prevents its switching-off by ecdysone at adult commitment is still unclear (Zhou, 2002).
These studies demonstrate for the first time that by the misexpression of a single transcription factor of the ecdysone cascade, the Br-Z1 isoform, one can redirect cells undergoing either larval or adult differentiation into a pupal developmental program. They also provide the first molecular basis for the 'status quo' action of JH on the pupal-adult transformation, by showing that JH causes the re-induction of Br expression and consequently re-expression of the pupal program during the molt (Zhou, 2002).
The ecdysis behavioral sequence in insects is a classic fixed action pattern (FAP) initiated by hormonal signaling. Ecdysis triggering hormones (ETHs) release the FAP through direct actions on the CNS. This study presents evidence implicating two groups of central ETH receptor (ETHR) neurons in scheduling the first two steps of the FAP: kinin (aka drosokinin, leucokinin) neurons regulate pre-ecdysis behavior and CAMB neurons (CCAP, AstCC, MIP, and Bursicon) initiate the switch to ecdysis behavior. Ablation of kinin neurons or altering levels of ETH receptor (ETHR) expression in these neurons modifies timing and intensity of pre-ecdysis behavior. Cell ablation or ETHR knockdown in CAMB neurons delays the switch to ecdysis, whereas overexpression of ETHR or expression of pertussis toxin in these neurons accelerates timing of the switch. Calcium dynamics in kinin neurons are temporally aligned with pre-ecdysis behavior, whereas activity of CAMB neurons coincides with the switch from pre-ecdysis to ecdysis behavior. Activation of CCAP or CAMB neurons through temperature-sensitive TRPM8 gating is sufficient to trigger ecdysis behavior. These findings demonstrate that kinin and CAMB neurons are direct targets of ETH and play critical roles in scheduling successive behavioral steps in the ecdysis FAP. Moreover, temporal organization of the FAP is likely a function of ETH receptor density in target neurons (Kim, 2015).
Innate behaviors are stereotypic patterns of movement inherited from birth that require no prior experience for proper execution. Among such behaviors are fixed action patterns that, once initiated, run to completion independent of sensory inputs. Examples include courtship rituals, aggression displays, and ecdysis. Ecdysis represents a 'chemically-coded' behavioral sequence triggered by peptidergic ecdysis triggering hormones (ETH), which orchestrate a downstream peptidergic cascade leading to sequential activation of central pattern generators underlying patterned motor activity. The term Fixed Action Pattern (FAP) has fallen into disuse, since innate behaviors generally exhibit considerable plasticity. However the invariant nature of the ecdysis behavioral sequence makes it a clear example a classic FAP. In depth analysis of ecdysis behavior may provide a more thorough understanding of how hormones assemble and regulate behavioral circuitry in the brain, in particular circuits that operate sequentially (Kim, 2015).
ETHs are released by peripheral peptidergic Inka cells in response to declining levels of the steroid hormone 20-hydroxyecdysone. Presence of Inka cells in more than 40 species of arthropods, along with the sequence similarity of ETH peptides in diverse insect groups, suggests that ETH signaling is highly conserved in insects. Identification of the Ecdysis Triggering Hormone receptor (ETHR) gene in Drosophila melanogaster (Park, 2002) enabled elucidation of a complex downstream signaling cascade triggered by ETH. The ETHR gene encodes two functionally distinct subtypes of G protein coupled receptors (ETHR-A and -B) through alternative splicing. The presence of two ETH receptor subtypes has been observed in all insect species thus far examined (Roller, 2010). The two receptor subtypes show differences in ligand sensitivity and specificity and are expressed in separate populations of central neurons, suggesting that they have distinctive roles in ETH signaling (Kim, 2015).
A diversity of ETHR neurons in the moth Manduca sexta and fruitfly Drosophila melanogaster has been identified (Kim, 2006a; Kim, 2006b). One of the most striking properties of ETHR-A neurons is that they are virtually all peptidergic and conserved across insect orders. Groups of ETHR-A 'peptidergic ensembles' express a range of different neuropeptides, including kinins, diuretic hormone (DH), eclosion hormone (EH), FMRFamide, crustacean cardioactive peptide (CCAP), myoinhibitory peptides (MIPs), bursicon (burs and pburs), neuropeptide F (NPF), and short neuropeptide F (sNPF). It is hypothesized that released ETH acts directly on the CNS to activate these peptidergic ensembles for control of specific central pattern generator circuits that elicit stereotyped ecdysis behaviors. However evidence for direct actions of ETH on these target ensembles is yet to be reported (Kim, 2015).
Likely functions of certain ETHR-A peptidergic ensembles in Manduca have been inferred from pharmacological manipulations (Kim, 2006b). For example, serially homologous L3/4 neurons of abdominal ganglia in Manduca express a cocktail of kinins and diuretic hormones; exposure of the isolated CNS to these peptides elicits a fictive pre-ecdysis I-like motor pattern. Similarly, the IN704 peptidergic ensemble that co-expresses CCAP and MIPs is implicated in initiation of ecdysis behavior, since co-application of these two peptides elicits fictive ecdysis behavior. Homologous peptidergic ensembles in Drosophila exhibit characteristic patterns and time courses of calcium mobilization indicative of electrical activity coincident with successive steps in the ecdysis FAP (Kim, 2006a). Of particular interest are observations that bursicon, a hormone co-expressed in a subset of CCAP neurons, is required for ecdysis behavior (Kim, 2015).
This study tested hypotheses that two central peptidergic ensembles—kinin neurons and a subset of CCAP neurons (CAMB) that co-express CCAP, Allatostatin CC, Myoinhibitory peptide, and Bursicon—are direct targets of ETH and schedule pre-ecdysis and ecdysis behavior components of the ecdysis FAP, respectively in the fruit fly Drosophila. It was shown further that manipulation of ETHR expression levels and signal transduction specifically in these ensembles influences scheduling of the FAP. Finally, possible mechanisms are described underlying timing of the switch from pre-ecdysis to ecdysis behavior and a model is proposed to explain mechanistically how these behaviors are sequentially activated (Kim, 2015).
The aims of this study were to implicate identified ETHR neuronal ensembles with specific steps in the ecdysis FAP. Through genetic manipulation of all known ETHR-A central neuron ensembles and subsets thereof, kinin and CAMB neurons were implicated in scheduling of pre-ecdysis and ecdysis, respectively. The current findings confirm that these ensembles are targeted directly by ETH. A key observation is timing of calcium mobilization in kinin and CAMB ensembles following ETH application: kinin neurons mobilize calcium within minutes, while activity in CAMB neurons is delayed. CAMB neurons mobilize calcium only after kinin neuron activity ceases, some 10 minutes later. These temporal patterns of cellular activity correspond well with those of pre-ecdysis and ecdysis behaviors observed in vivo (Kim, 2015).
The findings demonstrate that the kinin peptide ensemble is necessary for proper scheduling of pre-ecdysis behavior, if not itself sufficient to elicit it. Kinin cell ablation abolishes pre-ecdysis behavior in a significant percentage (25%) of animals. The remaining 75% of individuals showed highly variable pre-ecdysis duration, ranging from 3-22 minutes, whereas duration of this behavior in control animals is tightly regulated at 9.1 ± 0.9 min. Furthermore, a 30% reduction in Leukokinin receptor expression, caused by a piggyBac-element insertion into the promoter region of the gene, also disrupts fidelity of pre-ecdysis regulation; this phenotype is rescued by precise excision of the piggyBac insertion. Finally, RNA silencing of the kinin receptor in peripheral neurons using the Peb-Gal4 driver leads to reduced intensity of the behavior and greatly increases variability of pre-ecdysis duration. This is the first report demonstrating that kinin signaling affects scheduling of the ecdysis behavioral sequence via actions on peripheral neurons. Pan-neuronal silencing of kinin receptors also disrupted pre-ecdysis scheduling, but to a lesser extent (Kim, 2015).
Manipulation of ETHR expression levels in kinin neurons also alters scheduling of pre-ecdysis behavior significantly, confirming that these neurons are targeted directly by ETH and that they play an important role in pre-ecdysis regulation. It was reasoned that knockdown of receptor expression in these neurons would lead to a lower density of ETHR in the plasma membrane, thereby reducing sensitivity to ETH and delaying onset of pre-ecdysis. These experimental results demonstrated a reduction of pre-ecdysis duration. This reduction is attributed to a delay in pre-ecdysis onset, brought about by the need for higher ETH levels for neuronal activation. Since timing of the switch to ecdysis (controlled by CAMB neurons; see below) is unaffected, pre-ecdysis duration is shortened. On the other hand, overexpression of ETHR in kinin neurons led to prolongation of pre-ecdysis duration. Following the same reasoning, this would result from premature kinin neuron activation attributable to higher sensitivity to rising ETH levels and overall longer pre-ecdysis (Kim, 2015).
Kinins were identified originally using bioassays for myotropic and diuretic functions and they have well-known actions on muscle and transport activity in epithelia. More recent studies demonstrated diverse functional roles for kinin signaling, including feeding, olfaction, and locomotory behavior. Previous works demonstrated ETHR-A expression in kinin neurons of both Drosophila and Manduca, implicating them as direct targets of ETH. Imaging studies have shown that abdominal kinin neurons in fly larvae exhibit periodic calcium oscillations under normal conditions and are involved in turning behavior. These kinin neurons project to a terminal plexus in close association to kinin receptors, suggesting it functions as a site of peptide release. Interestingly, this same ensemble of kinin neurons in the pre-pupal preparation used here showed no such periodic activity, but instead exhibited synchronized calcium oscillations activity following exposure to ETH. This difference could be unique to the pharate stage (i.e., preceding ecdysis) animal, during which insects generally are unresponsive to external stimuli. In imaging studies, ETH-induced calcium dynamics were observed to be initiated from the terminal plexus region and subsequently move anteriorly to the cell bodies. These observations suggest that this plexus serves critical functions in both sending and receiving signals. Evidence presented in this study for regulation of pre-ecdysis behavior by kinin neurons demonstrates a new function for this peptide in Drosophila, which is reinforced by previous observations in Manduca that application of kinin causes a fictive pre-ecdysis motor pattern in the isolated CNS (Kim, 2015).
How do kinin neurons function in the promotion of pre-ecdysis behavior? While manipulation of kinin signaling clearly affects behavioral intensity and duration, this study was unsuccessful in initiating pre-ecdysis through temperature-dependent activation of kinin neurons expressing either TRPM8 and TRPA1. It is concluded that, while kinin functions as a modulatory influence necessary for proper scheduling of pre-ecdysis behavior, other as yet unidentified signals are necessary for behavioral initiation (Kim, 2015).
This study has demonstrated that CAMB neurons are both necessary and sufficient for the switch from pre-ecdysis to ecdysis behavior. This conclusion rests on results from a combination of experiments. First, CAMB cell ablation abolishes the switch to ecdysis, suggesting these neurons are necessary for the switch. Failure of ecdysis initiation is attributable to bursicon deficiency, since a previous report showed clearly that expression of the bursicon gene is required for initiation of pupal ecdysis. Calcium mobilization in CAMB neurons is delayed for ~10 min after onset of activity in kinin neurons, which fits well with the ~10 min delay before appearance of ecdysis behavior following onset of pre-ecdysis behavior observed in vivo. Altered levels of ETHR expression in CAMB neurons clearly affects timing of the switch to ecdysis behavior: receptor knockdown delays the switch, whereas overexpression accelerates it. In vitro experiments confirm that altered ETHR expression levels affect timing of calcium mobilization in CAMB neurons in register with changes in behavioral timing (Kim, 2015).
Finally, this study showed that activation of CAMB neurons through temperature-sensitive TRPM8 expression initiates ecdysis behavior in vivo. Thus, CAMB neurons are both necessary and sufficient for the switch to ecdysis behavior. However, activity in CAMB neurons alone does not result in robust ecdysis behavior. Expression of ecdysis behavior with parameters corresponding to that observed in wild-type flies requires activation of the entire CCAP ensemble (Kim, 2015).
It is interesting that, in all TRPM8 activation experiments, removal of the temperature stimulus led to re-capitulation of the entire ecdysis FAP. This might be explained by positive feedback influences on the Inka cell to release ETH, possibly via EH neurons. Alternatively, the ecdysis motor circuit may exert negative feedback on the pre-ecdysis circuit, which when removed, causes a post-inhibitory rebound leading to activation of the pre-ecdysis circuit and the entire FAP. Attempts to demonstrate such negative feedback were inconclusive in this study (Kim, 2015).
CAMB neurons express a combination of CCAP, Ast-CC, MIP, and bursicon. In Manduca, application of a CCAP/MIP cocktail is sufficient to elicit fictive ecdysis behavior. It would be parsimonious to extrapolate this result to Drosophila, since both of these peptides are found in CAMB neurons. Nevertheless, in Drosophila it is clear that bursicon is a key signaling molecule necessary for ecdysis initiation. It remains to be demonstrated precisely how absence of the bursicon gene blocks the switch to ecdysis. It will be interesting to elucidate possible functional roles of co-expressed peptides in CAMB neurons CCAP, Ast-CC, MIP in activation of the motor circuitry encoding the ecdysis motor pattern (Kim, 2015).
How is timing of the switch to ecdysis determined? Since both kinin and CAMB ensembles express ETHR, one would expect ETH to activate both ensembles simultaneously. Several previous studies provide evidence for the role of descending inhibition from cephalic and thoracic ganglia in setting the delay in the switch to ecdysis behavior. This study shows that expression of pertussis toxin in CAMB neurons accelerates the switch to ecdysis, consistent with disinhibition of Gαi/o input(s). It is hypothesized that a balance of excitatory and inhibitory inputs to the CAMB neurons contributes to the delay in their activity, excitatory input coming from ETH via Gαq signaling and Gαi/o from an as yet unidentified transmitter descending from cephalic and/or thoracic ganglia. The finding that RNAi-knockdown of MIP neurons lying outside the CCAP ensemble accelerates the switch to ecdysis behavior suggests one such possible inhibitory input (Kim, 2015).
It is possible, if not likely that ETH drives both inhibitory and excitatory inputs to CAMB neurons, with ETHR-B-expressing inhibitory inputs preceding excitatory input. Such a scenario follows from the fact that sensitivity of Drosophila ETHR-B to ETH was shown to be ~450-fold higher than that of ETHR-A. Therefore, as ETH levels rise in the hemolymph, ETHR-B-expressing inhibitory neurons would be activated well before ETHR-A neurons. ETH would effectively inhibit CAMB neurons indirectly prior to direct excitation via ETHR-A activation (Kim, 2015).
Such a scenario pre-supposes that the EC50 values governing activation of ETH receptors determined previously from heterologous expression in mammalian CHO cells are valid in Drosophila neurons. Data presented in this study suggests this is so. The EC50 value for ETH1 against ETHR-A was found to be ~414 nM, while the EC50 for ETH2 was determined to be ~4.3 μM. This study applied a combination of ETH1 and ETH2, each at a concentration of 300 nM, to the isolated CNS and obtained a pattern of calcium dynamics in kinin neurons lasting for ~10 min, which matches the duration of pre-ecdysis behavior under natural conditions. Furthermore, the switch to ecdysis behavior occurs ~10 min after initiation of calcium mobilization in kinin neurons, which corresponds to timing of the switch to ecdysis behavior in vivo. Doubling concentrations of the ETH1/ETH2 cocktail reduced the duration of calcium dynamics in kinin neurons to 5.5 min and accelerated the switch to ecdysis behavior. These results make it likely that the relative sensitivities of ETHR-B and ETHR-A are as previously established and consequently activity in ETHR-B neurons would precede that of ETHR-A neurons (Kim, 2015).
This study has shown that altered levels of ETHR expression have significant consequences for timing of pre-ecdysis duration and timing of the ecdysis switch. These findings raise the possibility that scheduling of sequential steps in the ecdysis FAP may be a consequence of different sensitivities to the peptide ligand. In other words, delay in the switch to ecdysis could result from a lower density of ETHR in CAMB neurons, making them less sensitive to ETH. Possible differential sensitivity to ETH could be tested in variety of way, including assessing timing of responses to the ligand by acutely dissociated neurons and/or single cell PCR (Kim, 2015).
A mechanistic model is proposed to explain neural mechanisms underlying the Drosophila pupal ecdysis FAP (see A model depicting functional roles of kinin and CAMB neurons in scheduling of the ecdysis FAP). Principle players in orchestration of pre-ecdysis and ecdysis behaviors are the kinin and CAMB ETHR ensembles, respectively. As ETH levels rise in the hemolymph, ETHR-B neurons are activated due to their high sensitivity (EC50 ~ 1 nM). These neurons release inhibitory signals acting through Gαi/o to inhibit CAMB neurons. As ETH levels rise further, kinin neurons receive direct excitatory input from ETH signaling via ETHR-A and Gαq to mobilize calcium from intracellular stores, leading to electrical activity in these neurons. ETH acts simultaneously on CAMB neurons, but inhibition from ETHR-B neurons descending from anterior ganglia prevents them from becoming active. As inhibition wanes, CAMB neurons become active, initiating the switch to ecdysis behavior (Kim, 2015).
Neural networks are typically defined by their synaptic connectivity, yet synaptic wiring diagrams often provide limited insight into network function. This is due partly to the importance of non-synaptic communication by neuromodulators, which can dynamically reconfigure circuit activity to alter its output. This study systematically mapped the patterns of neuromodulatory connectivity in a network that governs a developmentally critical behavioral sequence in Drosophila. This sequence, which mediates pupal ecdysis, is governed by the serial release of several key factors, which act both somatically as hormones and within the brain as neuromodulators. By identifying and characterizing the functions of the neuronal targets of these factors, this study found that they define hierarchically organized layers of the network controlling the pupal ecdysis sequence: a modular input layer, an intermediate central pattern generating layer, and a motor output layer. Mapping neuromodulatory connections in this system thus defines the functional architecture of the network (Diao, 2017).
Using the Trojan exon method to selectively target populations of hormone receptor-expressing neurons for manipulation and monitoring of activity, this study investigated the neuromodulatory connectivity of the circuitry governing pupal ecdysis behavior in Drosophila. The sites of action of the neuromodulators ETH, Bursicon, and CCAP identify essential functional components of the network architecture, defining three hierarchically organized layers from the sites of hormonal initiation to the sites of motor neuron output. In addition, it was found that descending neuromodulatory signaling from the ETHR-expressing input layer not only governs the basic motor rhythms of the ecdysis sequence by modulating the intermediate CPG layer, but also modulates activity of the CCAP-R-expressing motor neurons of the output layerd. Neuromodulators thus act broadly within, as well as across, network layers. The finding that the functional architecture of the ecdysis network can be decoded from its patterns of neuromodulatory connectivity provides further evidence that characterizing neuromodulatory connectomes is a valuable strategy in elucidating neural networks (Diao, 2017).
Major components of the pupal ecdysis circuitry are shared by the three motor programs
A schematic (ETHRB-expressing and non-CCAP/ETHRA neurons regulate Phase I) broadly augments existing models of the pupal ecdysis network, both by providing a more comprehensive description of the input layer than has previously been possible and by identifying the motor circuits on which this layer acts. A principal finding reported in this study is that the downstream targets of Bursicon and CCAP are shared components of the pupal ecdysis network and are used to generate all three motor rhythms. The results draw particular attention to the centrality of neurons that express the Bursicon receptor (Rk), which are absolutely required for all pupal ecdysis behavior. A role in central pattern generation is indicated both from the effects of their suppression, which eliminates all motor activity, and from their pattern of ETH1-induced Ca++ activity, which matches the phases of ecdysis behavior. The fact that ETH1-induced Ca++ activity is observed in the excised nervous system and thus in the absence of sensory feedback, demonstrates that it is centrally generated and further supports the identification of the VNC-Rk neurons as central pattern generators. Conclusive evidence that some or all VNC-Rk neurons participate in central pattern generation will require more precise observations and perturbations than those performed in this study, as will determining the functional roles of individual neurons. However, the preliminary observation that regions containing at most small numbers of VNC-Rk neurons exhibit activity that is phasically coupled to two or more motor patterns argues that the ecdysis circuitry includes multifunctional CPG neurons that express Rk and are subject to modulation by distinct input layer modules. Similar architectures have been described in other motor networks where two CPGs formed from overlapping pools of neurons can switch between activity states to generate distinct behaviors (Diao, 2017).
How input layer neurons modulate the pupal ecdysis CPG is exemplified by the control of Phase II by ETHRA/CCAP neurons. Direct activation of these neurons induces Phase II-like rhythmic activity in the VNC-Rk neurons, an observation that is easily explained if Bursicon secreted from ETHRA/CCAP neurons shifts the mode of activity of the VNC-Rk CPG. This mechanism is consistent with the neuromodulatory control of CPGs described in numerous other systems and accounts for the long-standing observation that CCAP- and Bursicon-expressing neurons are important for pupal ecdysis, including Phase II ('ecdysis') initiation and Phase I ('pre-ecdysis') termination. The CCAP- and Bursicon-expressing neurons are known to express additional neuropeptides, including Myoinhibitory Peptides and Allatostatin C, and it is likely that these neuromodulators also play a role in regulating these phases. The mixed activity patterns that define the transition from Phase II to Phase III ('post-ecdysis') are also readily interpreted as a period of bistability in which CPG modes transiently alternate, perhaps as Bursicon and/or other co-released neuromodulator concentrations fall (Diao, 2017).
In addition to neurons that switch CPG activity from Phase I to Phase II, the input layer must also contain neurons that initiate pupal ecdysis by inducing Phase I. The search for such neurons has focused primarily on those that express ETHRA, but no components of this group have yet been identified that are required for ecdysis initiation. To identify the ETH targets responsible for Phase I, ETHR-expressing neurons were systematically parsed into three, nearly mutually exclusive subsets that together cover the entire input layer. The results indicate that the largely uncharacterized neurons that express the B-isoform of ETHR are required to initiate Phase I, and that the non-CCAP/ETHRA neurons are important for maintaining that phase (Diao, 2017).
The essential role of ETHRB-expressing neurons in Phase I initiation is consistent with the significantly higher affinity for ETH peptides of ETHRB compared with ETHRA. ETHRB-expressing neurons may thus initiate Phase I by responding to rising titers of ETH earlier than neurons expressing ETHRA. How they regulate the VNC-Rk CPG neurons remains to be determined, but their mechanism of action appears to be different from that of the ETHRA/CCAP neurons insofar as the Phase I motor program cannot be evoked by TrpA1-mediated activation. It could be that this manipulation fails to induce the correct pattern of activity in ETHRB-expressing neurons. Preliminary imaging results show that ETHRB-expressing neurons respond to ETH1 with oscillatory activity, and it is possible that these neurons directly couple to the Rk-expressing neurons through synaptic or electrical contacts and participate in generating Phase I behavior. However, further characterization of the activity of both the ETHRB- and non-CCAP/ETHRA neurons will be required to determine how they modulate VNC-Rk CPG activity (Diao, 2017).
Two input layer neurons that are common to the ETHRB- and non-CCAP/ETHRA groups express the major ecdysis neuromodulator, EH (Diao, 2015). The EH-expressing neurons, which are among the few cells to express both ETHRA and ETHRB, respond to ETH1 application at the onset of Phase II, and evidence from other insects indicates that EH targets CCAP-expressing neurons. EH is thus thought to be responsible for the release of CCAP and Bursicon, but this has not yet been verified in Drosophila where the EH receptor has yet to be identified. It was thus not possible to target EH receptor-expressing neurons in this study, but the identity and function of such neurons is likely to be critical to understanding the progression of the ecdysis sequence (Diao, 2017).
In general, it is worth noting that the neuromodulators regulating the ecdysis sequence are of the type called 'extrinsic,' because they are released from neurons that do not function in the circuits upon which they act. Extrinsic neuromodulatory neurons, however, must be components of the broader neural networks that generate behaviors, and the mechanisms that organize their activities are only beginning to be understood. In some cases, these mechanisms are surprising. For example, the neuromodulatory connections between neurons that govern two foraging states in C. elegans are orthogonal to the sensory-to-motor synaptic connections between these neurons, which are not involved in the state decision. There are currently few studies that jointly examine patterns of neuromodulatory and synaptic connectivity, and to understand how extrinsic neuromodulatory neurons integrate into the broader networks in which they function more examples of such networks are required. Elucidating the interactions of neurons in the input layer of the ecdysis network (in addition to interactions of the input layer with neurons in other layers) should provide insight into this general problem (Diao, 2017).
The finding that the motor output of the pupal ecdysis network is mediated by neurons that express the CCAP-R provides insight into the hitherto poorly understood mechanism of action of CCAP. This neuropeptide plays critical roles in the ecdysis of other insects, but genetic data demonstrate that in Drosophila it plays a subsidiary role to Bursicon, acting synergistically with that hormone to render pupal ecdysis more robust. The current results indicate that it does so by acting on motor neurons, and because CCAP is co-released with Bursicon from the ETHRA/CCAP neurons to govern the CPG transition at Phase II, this suggests a role for feed-forward signaling in the pupal ecdysis circuit (Diao, 2017).
Neuromodulatory feedforward pathways have been previously described and appear to be a common motif in motor network architectures. Feedforward loops of the type posited here for Bursicon and CCAP may be important in adjusting the coupling between Rk-expressing CPG neurons and their downstream motor neuron targets during Phase II. Compensatory adjustments in CPG, motor neuron and muscle activity by a single neuropeptide released from two different nodes in a feedforward loop have been described in the Aplysia feeding network where they guarantee stability of network output. Coordinating CPG activity with motor neuron activity may be particularly important for multifunctional CPGs, in which individual neurons participate in multiple motor patterns, as for example, in the leech swim/crawl network in which multifunctional neurons fire in phase with the contraction of one muscle group during swimming, but not necessarily during crawling (Diao, 2017).
The architecture of the pupal ecdysis network revealed in this study is similar to that of other motor circuits, such as those governing locomotion, feeding, and breathing in which higher order neurons modulate the activity of core CPGs to generate varied motor patterns. What is striking about neuromodulator action in the ecdysis circuit is its broad scope. ETH acts throughout the input layer to control different phases of pupal ecdysis behavior; Bursicon similarly regulates a large and essential set of neurons constituting the ecdysis CPG; and CCAP acts on many motor neurons necessary for generating the rhythmic ecdysis movements. The observation that Bursicon and CCAP signal from the input layer speaks to an organizational logic in which the ecdysial neuromodulators function together to provide coherence to the operation of the pupal ecdysis network by acting both within each hierarchical layer and by acting coordinately across layers. This organization is consistent with a generalized role for neuromodulatory systems in organizing neural activity to generate behavior (Diao, 2017).
The results also support the rationale of mapping neuromodulatory pathways as a strategy for identifying essential network circuits and their functional organization. It is worth noting that this mapping of the pupal ecdysis network was done without reference to patterns of synaptic connectivity. Synaptic connectomes have proved difficult to interpret, in part due to their dense interconnectivity. If, as has been previously emphasized, this interconnectivity reflects the multifunctionality of the underlying networks, and if the functional configuration of a network at any given time is determined by where and how neuromodulators are acting on its components, then patterns of neuromodulatory connectivity may provide a necessary complement to synaptic maps to render them interpretable. A key challenge will lie in identifying which neuromodulator systems play critical roles in establishing a network's output, but as the work here demonstrates, when these are known, the neuromodulatory connections can deliver substantial insight into how a neural network is organized (Diao, 2017).
This study has focused on the mushroom bodies (MB) of Drosophila to determine how the larval circuits are formed and then transformed into those of the adult at metamorphosis. The adult MB has a core of thousands of Kenyon neurons; axons of the early-born g class form a medial lobe and those from later-born a'b' and ab classes form both medial and vertical lobes. The larva, however, hatches with only g neurons and forms a vertical lobe 'facsimile' using larval-specific axon branches from its g neurons. Computations by the MB involves MB input (MBINs) and output (MBONs) neurons that divide the lobes into discrete compartments. The larva has 10 such compartments while the adult MB has 16. This study determined the fates of 28 of the 32 types of MBONs and MBINs that define the 10 larval compartments. Seven larval compartments are eventually incorporated into the adult MB; four of their larval MBINs die, while 12 MBINs/MBONs continue into the adult MB although with some compartment shifting. The remaining three larval compartments are larval specific, and their MBIN/MBONs trans-differentiate at metamorphosis, leaving the MB and joining other adult brain circuits. With the loss of the larval vertical lobe facsimile, the adult vertical lobes, are made de novo at metamorphosis, and their MBONs/MBINs are recruited from the pool of adult-specific cells. The combination of cell death, compartment shifting, trans-differentiation, and recruitment of new neurons result in no larval MBIN-MBON connections persisting through metamorphosis. At this simple level, then, no anatomical substrate was found for a memory trace persisting from larva to adult. For the neurons that trans-differentiate, the data suggest that their adult phenotypes are in line with their evolutionarily ancestral roles while their larval phenotypes are derived adaptations for the larval stage. These cells arise primarily within lineages that also produce permanent MBINs and MBONs, suggesting that larval specifying factors may allow information related to birth-order or sibling identity to be interpreted in a modified manner in these neurons to cause them to adopt a modified, larval phenotype. The loss of such factors at metamorphosis, though, would then allow these cells to adopt their ancestral phenotype in the adult system (Truman, 2023).
Direct developing insects like crickets and grasshoppers produce a species-typical body form during embryogenesis and hatch as a miniature version of the adult but lack wings and genitalia. Holometabolous insects, which have complete metamorphosis, have modified their embryogenesis to produce a simplified larval body instead. In the larval body, embryonic fields, which had been fully utilized to generate their adult-like structures, are only partially patterned, with the patterned portion making the larval structure and the remainder preserved through larval growth to become imaginal discs or primordia. The imaginal primordia, along with some larval cells, combine to construct the species-typical body form at metamorphosis (Truman, 2023).
Direct development also results in a hatchling possessing an essentially mature, but miniature brain, which is used for both the nymphal and adult stages. The evolution of the larva, though, altered brain development to make a modified, simpler brain appropriate for the sensory and motor demands of the larva. This larval brain, though, is not discarded at the end of larval growth and a new one made from scratch. Most larval neurons persist and some, like the interneurons mediating backwards locomotion, have similar functions in both larva and adult, but, as is shown in this study, the maintenance of neuronal function from larva to adult is not always the case. At metamorphosis, recycled larval cells are combined with adult-specific neurons to make the nervous system of the adult (Truman, 2023).
Although the focus on nervous system metamorphosis is usually on the postembryonic transformation of the larval brain into that of the adult, complementary changes had to have occurred during embryogenesis to generate the modified larval brain. One key embryonic change involved neurogenesis. The central brain and ventral nerve cord (VNC) of insects arise from a fixed number of neuronal stem cells (neuroblasts [NBs]), with about 100 per brain hemisphere and about 30 per segmental hemineuropil. Each NB makes a characteristic set of neurons in a defined temporal order. The sets of NBs in the brain and VNC, though, are highly conserved and were established well before the evolution of the larva and complete metamorphosis. Moreover, these conserved sets of NBs produce the neurons for both the larval and adult CNS. The duration of embryonic neurogenesis, however, differs in the two groups. Insects with direct development, like grasshoppers, produce all the neurons of the central brain and VNC during embryogenesis and the hatchling possesses all the neurons of the adult (except for expansion of the Kenyon cell population,). In insects with complete metamorphosis, by contrast, selection for the rapid formation of a larval stage required a premature arrest of neurogenesis, resulting in many fewer neurons for the larval brain. In Drosophila, for example, most brain and thoracic NBs produce only 10–15% of their respective progeny by hatching and the remainder are made during a second neurogenic period late in larval growth. Since essentially all their neurons are born during embryogenesis, the hatchlings of direct developing insects have brain circuits that include neurons generated during early, intermediate, and late phases of their neuroblasts' lineages. The larvae of metamorphic insects, by contrast, not only have fewer neurons to make their brain, but these neurons include only those with early-born fates (Truman, 2023).
A second key change involves the phenotypes of the neurons that are made during embryogenesis. The neurons of direct developing insects acquire their mature phenotype by the time of hatching and their anatomy and connections change very little through nymphal growth and adulthood. By contrast, in holometabolous insects, the form and function of many larval neurons are radically different from their adult form and function. At metamorphosis, they lose their larval specializations and finally acquire their mature, adult phenotypes (Truman, 2023).
The analysis of development and metamorphosis of complex neuropils can provide important insights into the mechanisms underlying the formation of the larva and its subsequent metamorphosis. This study has focused on the larval and adult circuitry of the mushroom bodies (MBs). In both stages, the MBs associate odors with either rewards or punishments and adjust the animal's future behavior accordingly. The circuitry of the MB is known at the EM level for both the larva and adult. The core of the MB is a set of hundreds (larva) to thousands (adult) of small neurons called Kenyon cells. Their dendrites project into calyx neuropil, which receives primarily olfactory input via antennal lobe projection neurons, and into accessory calyx neuropils that receive visual and thermal information. The bundled axons of the Kenyon cells extend down the peduncle and into the vertical and medial lobes. The mature, adult MB has three major classes of Kenyon cells: the γ neurons made during embryogenesis and early larval life, the α'β' cells generated late in larval life, and the αβneurons born through early and mid-metamorphosis. There is considerable complexity within each of these major Kenyon cell classe, but overall, their axons form three medial lobes (γ, β', β) and two vertical lobes (α, α'. Unlike the adult, the MB of the larva contains only modified γ Kenyon cells whose axons form a vertical and a medial lobe (see The organization and development of the larval and adult mushroom bodies; Truman, 2023).
The MB receives flows of sensory information through projection neurons to the calyx, but this study focused on the sets of MB input neurons (MBINs) and output neurons (MBONs) that innervate the peduncle and lobes. In both larvae and adults, these neurons divide the lobes into non-overlapping compartments that have a common microcircuit motif. Each compartment is defined by the axonal tuft of an aminergic input cell that synapses onto Kenyon cell axons and onto a dedicated MBON(s). The Kenyon cell axons synapse onto each other and the MBONs but also feed back onto the MBINs. The majority of the MBINs are dopaminergic neurons (DANs) but a few are octopaminergic neurons (OANs). Most DANs come from two clusters, the protocerebral anterior medial (PAM) cluster, which primarily encodes reward, and the protocerebral posterior lateral 1 (PPL1) cluster, which mainly encodes punishment. Depending on their compartment, the MBONs are cholinergic, GABAergic, or glutaminergic. Interestingly, stimulation of the MBONs from PPL1-supplied compartments generally evokes approach behavior while stimulation of those from PAM supplied compartments evokes avoidance. Consequently, pairing punishment with a particular odor reduces the drive on MBONs that promote attraction. Behavior is therefore guided by a balance of avoidance versus attractive influences and the inhibition of neurons mediating one behavior then favors the opposite behavioral state. The functions of some compartments, though, are complex because of extensive interconnections amongst MBONs and feedback from MBONs back to various MBINs (Truman, 2023).
While serving similar functions of mediating associative learning, the larval and adult MB differ in a couple of ways. The larval MB has 10 compartments plus the calyx, while the adult has 16. The larval structure lacks the α'β' and αβ neurons that form the adult system's vertical lobes. The larval γ neurons, though, have larval-specific vertical axon branches that form the core of a larval vertical lobe. Focus has been placed on the MBINs and MBONs that establish the 10 larval compartments. The metamorphic fates of about 80% of these neurons has been determined. Depending on the fate of their compartment, some larval neurons remain with the MB, others reprogram to join other adult circuits, and still others die. The persistence of MBON to MBIN connections would have been the simplest way that a larval memory trace could be carried through metamorphosis. However, it has been found that the diverse fates of the larval MB neurons plus the addition of new, adult-specific MBINs and MBONs to the compartments result in no MBIN-MBON pairings that survive from larva to adult (Truman, 2023).
Some neurons like the GABAergic APL have a unique and characteristic morphology that allows them to be readily identified in both larval and adult stages and the same name has been used for both stages. The vast majority of MBINs and MBONs, however, have been given different names in the larva versus the adult, and this study determines their correspondence for the first time. Where useful in the 'Discussion,' their larval and adult names will be combined. For example, since larval MBON-d1 becomes adult MBON 11, it will be referred to as MBON-d1/MBON 11 (Truman, 2023).
The evolution of a metamorphic life history in insects required two changes: first, the modification of embryogenesis to produce a larval body; and second, metamorphosis itself –the transformation of that larval body into the insect's mature form. The focus of this study has been on the second process of transforming the larval MB into its adult form, but the results also provide insight into the first issue, i.e., how embryonic development may have been altered to form a specialized larval MB (Truman, 2023).
Focus has been placed on the metamorphic fates of the MBINs and MBONs that make the microcircuits that divide the MB lobes into their computational compartments (see Stability and changes in mushroom body (MB) compartments during metamorphosis). Three compartments are specialized for the larval stage and not carried into the adult. For the two distal vertical lobe compartments, UVL and IVL, their γ axon core degenerates and two of their MBONs (MBON-e2 and MBON-f2) switch to adult medial lobe compartments while their remaining MBONs and MBINs shift to adult circuits outside of the MB. The larval vertical lobe is then replaced with new, adult vertical lobes formed from α' and α axon cores and postembryonic-born MBINs and MBONs. There is, however, no cellular continuity between the larval vertical lobe and mature vertical lobes of the adult MB. The larval structure is thought as a vertical lobe 'facsimile'; a larval innovation to deal with the lack of the Kenyon cells and extrinsic neurons that typically make the vertical lobes (Truman, 2023).
The larva has two peduncle compartments, IP and LP, as compared to the single compartment (PED) of the adult. The larval LP compartment with some of its MBINs becomes incorporated into the adult PED compartment, but the larval IP compartment is lost and not replaced. Its two MBINs and at least two of its three MBONs leave the larval MB and become parts of adult non-MB circuits. The IP compartment is unusual because of its isolation from the other MB microcircuits. The latter are highly interconnected by one- or two-step connections from the MBONs of one compartment to the MBIN of others. The IP MBINs, however, receive no MBON feedback from the other compartments. Likewise, the IP MBONs provide the least amount of crosstalk to the other larval compartments. Its isolation suggests that the IP is involved in a type of learning distinct from that handled by the rest of the larval MB. This may involve temperature-based learning since the greatest input from the small number of thermosensory Kenyon cells is onto MBON-b3. Since no corresponding compartment in the adult MB has been, the type of learning that IP mediates may be restricted to the larva (Truman, 2023).
The remaining seven larval compartments are incorporated into adult MB compartments, especially those containing γ neuron axons (see Stability and changes in mushroom body (MB) compartments during metamorphosis). Their larval MBINs and MBONs typically also function in the mature MB, although the four PAM MBINs that project to the distal-most larval compartments (SHA, UT, IT, and LT) die at the end of the larval stage. The larval MBONs from these distal compartments, though, survive and innervate compartments at similar positions along the adult medial lobes, although some shift their dendrites from γ into β or β' compartments. For example, the larval SHA compartment corresponds to the adult γ3 compartment; of its two MBON-h's, one (MBON 08) innervates the adult γ3 compartment while the other (MBON 09) has dendrites in β'1 as well as γ3. The MBONs of the larval distal 'toes' (the UT, IT, and LT compartments) distribute their dendrites amongst the adult distal medial lobe compartments (γ4, γ5, β2, and β'2). The basal larval compartments, LP, LA, and LVL, correspond to adult compartments PED, γ1, and γ2, respectively. Most of their MBINs and MBONs stay within this set of compartments but there is some shuffling amongst them. The general pattern is that larval MBINs and MBONs that supply the seven larval compartments that are incorporated into the adult MB continue to function in the adult MB, while those that innervate the three larval-specific compartments retract from the MB and join non-MB circuits of the adult brain (Truman, 2023).
From the perspective of the mature, adult MB, 10 of its 16 compartments have axon cores from the postembryonic born α'β' or αβ Kenyon cells. The α and α' compartments are supplied almost exclusively by postembryonic-born MBINs and MBONs. By contrast, the β and β' compartments show a mixed picture: their input is provided exclusively by postembryonic-born PAM neurons, while their known outputs are through embryonic-born neurons from the larval MB (Truman, 2023).
The input and output transmitters associated with MB compartments of the larval and adult systems are similar but not identical. For the MBINs, the calyx receives octopaminergic input from the same neurons in the two stages. The adult calyx also receives a sparse serotonin input from a remodeled CSD neuron that projects from the contralateral antennal lobe. The compartments of the larval lobe system are primarily supplied by dopaminergic neurons, but the UVL and LVL compartments also have octopaminergic input. The octopamine input to the lobes is reduced in the adult MB, though, because OAN-e1/PPL1-SMP retracts completely from the larval UVL compartment and innervates the superior medial protocerebrum in the adult, and OAN-g1/OA-VPM3 reduces its MB input to a sparse innervation of the adult γ1 compartment (Truman, 2023).
The PPL1 and PAM clusters provide the major dopamine input to the lobes. The number of PPL1 neurons is about the same in the larval and adult stages, but those innervating vertical lobe compartments differ in the two stages. In contrast, the PAM cluster innervation of the medial lobes is dramatically increased as the four neurons in the larva are replaced by about 150 neurons in the adult. Appetitive conditioning in the adult is complex with different medial lobe compartments receiving PAM input from different brain regions and providing reward information based on diverse factors such as sugar sweetness, nutritional value, and water. Also, reproduction-related learning is also mediated through sets of PAM neurons (e.g., the PAM 01 ( = γ5) neurons). Even with only four PAM neurons, though, the larva shows selectivity in its reward learning. For example, inhibition of DAN-h1 interferes with the positive association and odor with a fructose reward, but not with either amino acid or low salt rewards (Truman, 2023).
An adult-specific, modulatory input to the MB circuitry is provided by the paired, larval-born DPM neurons that innervate all the compartments of the adult peduncle and lobes (but not the calyx). DPM neurons release serotonin and peptides produced from the amnesiac gene. These neurons are required for a delayed memory trace that appears in the MB about 30 min after training, and they participate in two forms of intermediate term memory, anesthesia-sensitive memory via the amnesiac gene, and anesthesia-resistant memory via serotonin and the radish gene. Anesthesia-resistant memory involving the radish gene also occurs in larvae. As the DPM neurons are postembryonic, it is unknown whether larvae use another modulatory neuron for anesthesia-resistant memory (Truman, 2023).
On the MBON side, the output from the calyx in both larva and adult is cholinergic. In the larval stage, both MBON-a1 and -a2 receive similar input from Kenyon cells and from OAN-a1 and -a2; MBON-a1 also excites MBON-a2 via axo-axonic synapses. MBON-a2/MBON 22 persists as the major cholinergic output from the adult calyx, while MBON-a1/MBON 29 shifts from the calyx to adult medial lobe compartments. MBON-c1/MBEN-CA is added to the adult calyx, but since its mature anatomy is unknown, it is not possibe to speculate on its function (Truman, 2023).
For the vertical lobe system, the output transmitters from the distal compartments of the adult medial and vertical lobes largely conform to those from the corresponding regions of the larval medial and vertical lobes. The adult α' and α (except α1) compartments have cholinergic output. The larval UVL compartment similarly has cholinergic output, but the transmitters of the MBONs from the IVL compartment are unknown (Truman, 2023).
The conservation of glutaminergic output of the distal medial lobe compartments between larva (UT, IT, and LT) and adult (γ4, γ5, β2, and β'2) comes from the same neurons being used in the two stages. The adult also has glutaminergic output from the α1 compartment provided by the two MBON 07s. These feedback on the α1 PAM neurons, thereby making a recurrent loop that is essential for appetitive long-term memory formation in the adult. It was not possible to. determine the developmental origin of MBON 07. In the larva, though, MBON-e2/MBON 03 provides an analogous glutaminergic output from the vertical lobes. It also feeds back onto its input neuron (OAN-e1), perhaps providing an analogous circuit to support long-term memory formation in the larva (Truman, 2023).
The compartments at the bases of the lobes and peduncle show some scrambling of neurotransmitter output through metamorphosis. The output from the larval LP compartment is cholinergic while the corresponding adult PED compartment has GABAergic output. The opposite shift is seen in the LVL ( = γ2) compartment, which has GABAergic output in the larva but cholinergic output in the adult. MBON-c1/MBEN-CA provides the cholinergic output from the larval LP compartment, but at metamorphosis it retracts from this compartment and dendrites from MBON-d1/MBON 11 invade the peduncle to provide the adult GABAergic output. The other compartmental shift in transmitter involves the larval LVL compartment; MBON-g1 and -g2 provide GABAergic output from this compartment, but at metamorphosis, they trans-differentiate to become central complex neurons. They are replaced by postembryonic-born MBON 12, which then provides cholinergic output for the adult γ2 and α'1 compartments. Thus, the compartmental shifts in output transmitters that occur at metamorphosis do not involve individual MBONs changing their transmitter. Rather, MBON recruitment, MBON loss, and MBONs shifting compartments combine to provide differences in transmitter landscapes in the two stages (Truman, 2023).
The examples of DAN-d1/PPL1 03 and DAN-c1/PPL1 01 show that shifting partners through metamorphosis can dramatically alter a neuron's function. In the larva, pairing of DAN-d1 stimulation with an odor induces short-term aversive conditioning, whereas a similar pairing with DAN-c1 does not. Their functions change in the adult, though, where DAN-c1/PPL1 01 becomes sufficient to induce short-term aversive conditioning to a paired odor, while DAN-d1/PPL1 03 becomes involved in higher levels of memory consolidation. The metamorphic changes in the functioning of these two MBINs comes from changing their MBON partners. In its larval form, DAN-d1/PPL1 03 works through MBON-d1/MBON 11 in establishing short-term aversive conditioning. This MBON functions similarly in the adult, but its adult input is provided by DAN-c1/PPL1 01 rather than DAN-d1/PPL1 03. DAN-d1/PPL1 03, in turn, instructs a new partner, MBON 12, a cholinergic, postembryonic-born neuron that provides feed-back excitation to DAN-d1/PPL1 03 and feeds across to a set of medial lobe MBONs whose activity promotes avoidance behavior, while their suppression promotes approach. These interactions provide a pathway in the adult to mediate memory re-consolidation and extinction (Truman, 2023).
The adult form of DAN-c1/PPL1 01 has the added complexity that the type of learning it supports reverses depending on the time of its activity relative to the paired odor stimulus. Its activation after presentation of the odor reinforces avoidance of the odor, but if the odor is presented after DAN-c1/PPL1 01 terminates its activity. The fly then shows a 'relief' response and the odor becomes attractive). It does not have such a function in the larva . In the larva, a similar time-dependent switch from appetitive to aversive learning is mediated through DAN-f1, a neuron that becomes incorporated into non-MB circuits in the adult (Truman, 2023).
Hence, the persisting MB neurons rearrange their connections at metamorphosis as some neurons are lost from the MB via trans-differentiation (e.g., MBON-c1) and other, adult-specific neurons are added (e.g., MBON 12). Such changes likely reflect ad hoc solutions that enable the construction of a larval circuit without needed late-born cell types by using other early-born neurons that display temporary phenotypes to make up for the missing cells. At metamorphosis, when the appropriate cell types are finally available, the temporary MB neurons trans-differentiate to evolutionarily ancestral phenotypes and the remaining neurons rewire into the adult circuit (Truman, 2023).
The connections between uni-compartmental MBINs and MBONs that are found in the larva , the adult, or both is summarized (see Fate of circuit connections in the mushroom body (MB) through metamorphosis). Stable MBIN-MBON pairing persists through metamorphosis only in the calyx neuropil, which contains the Kenyon cell dendrites. By contrast, the combination of compartment shuffling, trans-differentiation and neuronal death in the lobe compartments results in a lack of uni-compartmental MBIN to MBON pairings persisting from larva to adult (Truman, 2023).
Besides MBIN to MBON connections, the compartments of both the larva and the adult are highly interconnected, both by MBON-to-MBON connections and by feedback and feed-forward connections from MBONs back to MBINs. For MBON-to-MBON interactions, larval and adult connectivity data are available for seven of the MBONs that function in both circuits. There are 42 possible pair-wise connections amongst these cells. These MBONs are more highly interconnected in their adult configuration compared to their larval one: their adult configuration shows 13 connections (31% of possible connections), while their larval configuration has only 7 (17%). Importantly, only three of these connections (7%) are present in both larva and adult. This percentage is similar to the 5% predicted if the two stages were wired up independently at their respective frequencies. This low level of shared connections suggests that in both their larval and the adult configurations, the MBONs interconnect in a way that is best adapted to the respective life stage (Truman, 2023).
Experiments on aversive conditioning of Drosophila larvae suggested that the memory from larval training can persist through metamorphosis. This study found that within the MB lobe system, none of the MBIN-MBON pairings persist and persisting MBON-to-MBON connections are rare. At this level, anatomical findings do not identify any simple circuit elements that would support the persistence of an associative memory trace from larva to adult. Thus, a surviving memory trace would need to involve more complex anatomical pathways. However, this cannot be addressed in this study (Truman, 2023).
The failure to find anatomical support in Drosophila for persistence of a memory trace from larva to adult should not be generalized to other insects with a larval stage. There is evidence that associative learning in moth caterpillars and beetle grubs can carry through to the adult. Larvae of butterflies and beetles have an extended embryonic development compared to Drosophila, and they hatch with a more complex larval nervous system. Consequently, more of the neuron types needed to make their MB are available to these embryos, likely making these insects less dependent on appropriating other neurons to temporarily function in the larval MB. A higher number of MB neurons persisting from the larva to adult increases the likelihood that a memory trace could persist from one stage to the other (Truman, 2023).
Although there are examples of neurons that change their neuropeptides during postembryonic life, the current study did not find any neuron that changed its small molecule transmitter. The neurons did, though, show a great range of morphological changes. At one end of the spectrum were neurons like DAN-d1/PPL1 03 and MBON-j1/MBON 02 , whose larval and adult forms are very similar. At the other end of the spectrum are MBIN-b/PAL-OL and MBON-g/LALs-NO2i.b , which trans-differentiate into adult neurons that bear no similarity to their larval forms (Truman, 2023).
Neurons possessing the same form in both the larval and adult stages are like those of direct developing insects because they undergo their full developmental trajectory during embryogenesis and achieve their mature form at hatching. Other larval neurons have a morphology that appears to be based on pausing their developmental trajectory at an intermediate step and using this intermediate form as the basis for their larval morphology. The larval octopaminergic cells, OAN-a1 and -a2, fit this pattern. Their larval neurons stop at the MB calyx but in their adult form (OA-VUMa2), they extend beyond the calyx to form major arbors in the lateral horn neuropils. A similar strategy occurs for thalamic neurons in the developing mammalian visual system. These early-born neurons arrive at the cortical subplate prior to the birth of their granule cell targets in the visual cortex. They assume an intermediate phenotype, synapsing with the subplate neurons, but after the granule cells are born, they lose these temporary connections and grow into the cortex to find their final targets (Truman, 2023).
For other neurons, however, their larval form cannot be explained as a simple arrest along an ancestral developmental trajectory. The vertical axon branch of larval γ Kenyon cells is not seen as an intermediate stage in the development of γ neurons of direct developing insects such as crickets. The larval form of these neurons requires a developmental deviation that adds new features to adapt the neurons' morphology to the larval stage. The extreme version of adding larval-specific novelty is the radical change in cellular phenotype seen in trans-differentiating neurons like MBON-g and MBIN-b (Truman, 2023).
Cells that undergo trans-differentiation, like MBON-g/LALs-NO2i.b, show extensive pruning at the start of metamorphosis. Some neurons that have essentially the same morphology in larva and adult, like MBON-j1/MBON 04 and APL, also show extreme pruning. But others, like DAN-c1/PPL 01 or DAN-d1/PPL1 03, show only moderate arbor loss. This variation reflects the fact that while pruning is due to a cell autonomous developmental program triggered by the steroid ecdysone, its trajectory may be guided in some cells by local interactions with pre- or postsynaptic targets. The importance of local interactions was experimentally examined for the pruning of APL. Blocking ecdysone action in APL inhibits its pruning response. The selective inhibition of ecdysone action in γ Kenyon cells, the main synaptic partners of APL, similarly inhibits γ cell pruning but also that of the untreated APL. Thus, while steroid signaling is needed to activate the neuron's pruning program, the extent of neurite loss may depend on changes in synaptic partners. Similarly, for the MBINs, the larval LP and LA compartments retain γ neuron axons during pruning, and it is seen that their MBINs (DAN-c1 and DAN-d1) maintain most of their axonal tufts through the pruning period. By contrast, neurons of distal medial lobe compartments, which lose their γ neuron axons, prune extensively even though they grow back to a similar adult morphology (e.g., MBON j1/MBON 02 (Truman, 2023).
The MBINs and MBONs of the adult CNS are produced by 10–15% of the ~100 NBs that construct each brain hemisphere. Most of the neuron types that serve as temporary larval MB neurons are recruited from these same lineages. Although larval NBs and adult lineages have been mapped and described, the two maps have not been reconciled. Indeed, in most cases, it is not known exactly which embryonic brain NB produces which postembryonic lineage. The major embryonic and postembryonic lineages that produce MBINs and MBONs is described (see The lineage relationships of the major neuron types of the larval and adult mushroom bodies). The Kenyon cells are produced by the four MBps that begin dividing at mid-embryogenesis and only finish just before the emergence of the adult. Their earliest embryonic cells differ, but the four NBs produce identical lineages after they begin Kenyon cell production. All the other neuroblasts make a small number of neurons embryonically, but then arrest and subsequently resume cycling late in the first larval instar. Their small size during the dormant period makes them difficult to track through this transition (Truman, 2023).
Most MBINs come from the PPL1 and the PAM clusters. This study found that the generation of the adult PPL1 is split, with some members born in the embryo and initially functioning as larval MBINs, while others are born after neur ogenesis resumes in the larva (see The lineage relationships of the major neuron types of the larval and adult mushroom bodies). The embryonic neuroblast that makes these neurons is CPd2 or 3, and it appears to arrest in the midst of producing the PPL1 MBINs, a conclusion based on the observation that MBIN-c1 is born so late in embryogenesis that it is not yet incorporated into the MB circuitry at hatching. The neuroblast is called DL2 when it reactivates in the larva and shows a type II pattern of division. The first neurons that DL2 produces after it resumes dividing are the remaining PPL1 MBINs. The adult PPL1 neurons, therefore, appear to arise as a set of neurons that straddle the temporary arrest of the DL2 neuroblast. Those born in the embryo then function as MBINs in both the larva and the adult, while those born in the larva delay their maturation into MBINs until metamorphosis. Based on their clustering with the 'permanent MBINs,' the embryonic-born PPL1 neurons that serve as temporary larval MBINs (OAN-e1, MBIN-e2, and DAN-f1) arise in the same lineage but must be produced earlier in embryogenesis (Truman, 2023).
The adult PAM neurons are all born during the postembryonic period and are produced by two closely related lineages, CREa1A and CREa2A. The first few postembryonic neurons born in both lineages are not PAM neurons. However, both neuroblasts soon switch into a repetitive mode, in which the 'Notch-ON' daughter of each GMC becomes a PAM neuron, a pattern repeated through the next ~75 GMCs. The generation of this large neuronal class at the end of their lineages is consistent with the general pattern that neurons within a hemilineage become more similar as a neuroblast progresses deeper into its lineage (Truman, 2023).
Embryonic DAL CM1 and 2 are likely the embryonic version of CREa1A and 2A. One or the other makes four PAM neurons for use in the larva, but these subsequently die at metamorphosis. These temporary PAM neurons also seem to be born at the end of the embryonic neurogenic phase, because like MBIN-c1 described above, one of them, DAN-h1, is not yet incorporated in the MB circuitry at hatching. Interestingly, after the CREa2A neuroblast resumes dividing in the larva, its first Notch-ON daughter degenerates right after its birth. Towards the end of its embryonic phase, the CREa2A neuroblast is suggested to produce a set of GMCs, whose Notch-OFF daughter is required but whose Notch-ON daughter is 'unneeded' and fated to die. This pattern is carried through into the start of the postembryonic phase, as evident by the first postembryonic, Notch-ON daughter dying immediately after its birth. The Notch-ON daughters produced during embryogenesis, though, defer their deaths until metamorphosis and serve as temporary PAM neurons for the larva (Truman, 2023).
Members of various groups of MBONs are also related by lineage and by transmitter type. The adult has eight types of cholinergic MBONs that provide output from the α and α' compartments (25535793). Three come from the FLAa2 neuroblast and are produced at the beginning of its postembryonic phase. The remainder are produced within the postembryonic period by the DL1 neuroblast, another neuroblast that shows a type II pattern of division. Its divisions result in a series of GMCs that each divide to produce a cholinergic MBON and central complex neuron. Neither DL1 nor FLAa2 appear to contribute embryonic-born cells to the larval vertical lobe. The transmitters of the two IVL MBONs are not known, but MBON-e1, from the UVL compartment, is cholinergic. Curiously, it comes from the CPd2/3 group, a group that includes the PPL1 MBINs (Truman, 2023).
Besides the multicompartmental neuron, APL, the adult has four types of GABAergic MBONs . The β'1 compartment is innervated by eight MBON 10-type neurons. Their origin is not known although the large number of neurons in this group suggests that they are born during the postembryonic period. The remaining three types of GABAergic MBONs are embryonic-born and wholly or partially associated with γ compartments in both the larva and the adult. MBON-d1 comes from the DAL CM-1/2 group, the same group responsible for the PAM neurons. Of special interest, though, are MBONs 08 and 09 from the DAL-V2/3 group. These neurons have overlapping compartmental functions in the adult, but are identical in the larva, showing essentially the same synaptic connectivity within the MB. This lineage produces four, rather than two, GABAergic MBONs in the larva, but the additional two larval cells (MBON-g1 and -g2) trans-differentiate into adult central complex neurons. The BLVa3/4 group that produces APL also produces two larval-specific GABAergic neurons, MBON-b1 and -b2, that become lateral horn neurons in the adult (Truman, 2023).
The adult medial lobe compartments are supplied by seven types of glutamatergic MBONs. The origin of the adult MBON 07 is unknown, but the six other types are embryonic born, with the majority coming from the DAM-d1 lineage. These six types provide sufficient glutamatergic MBONs to cover larval medial lobe function, with an additional type leftover, MBON-e2/MBON 03, which is shifted to the larval vertical lobe. It provides glutamatergic output from the larval vertical lobe, perhaps analogous to that provided by MBON 07 from the adult α1 compartment (Truman, 2023).
Neuronal identity is established within a lineage by relative birth order of the GMCs (see The origin of spatial-temporal information used to determine neuronal phenotypes) and by symmetry-breaking to establish different fates of the two daughters of the GMC division. Relative birth order is encoded at the start of neurogenesis by the sequential expression of a series of transcription factors, Hunchback -> kruppel ->→ pdm -> castor, in the neuroblast as it divides (10993672 & 11525736). The transcription factor expressed at the time of division is passed into the GMC and then into her two daughter cells, thereby providing a record of relative birth order. In Drosophila, an embryonic neuroblast typically reaches castor expression by the time of its arrest late in embryogenesis and it typically resumes expressing castor when it reactivates in the larva. The phenotypes of the two siblings arising from the GMC division are established through Notch signaling, which results in a Notch-ON ('A') fate and a Notch-OFF ('B') fate. Successive cells sharing that same Notch state typically have similar properties, resulting in the neuroblast producing two hemilineages (A and B) of neurons of related form and function. The information on relative birth order and hemilineage status then acts through a set of terminal selector genes to produce a characteristic neuronal phenotype. These two mechanisms might be exploited to alter a neuron's phenotype for specialized use in the larva (Truman, 2023).
Information on birth order may underlie the changes seen in the PPL1 cluster. As described above, production of the set of neurons that become adult PPL1 MBINs spans the period of neuroblast arrest. It is speculated that this block of neurons expresses cas. In this scenario, their earlier born siblings that serve as temporary larval MBINs would likely express the previous gene in the series of temporal transcription factors, pdm. At metamorphosis the Pdm+ neurons remain aminergic but shift to targets outside of the core MB circuitry. The speculative scheme proposes that in the adult, Cas and Pdm work via overlapping sets of terminal selector genes: both establish a dopaminergic phenotype, but only Cas activates genes targeting the neuron to the MB. The intercalation of the larval stage, though, would involve an additional 'larval specification factor,' whose presence alters the actions of Cas and Pdm on the terminal selector genes. In the presence of this hypothetical factor, Pdm also supports targeting to the MB, thereby transforming the Pdm+ group into MBINs, which innervate the larval vertical lobe. The subsequent loss of the larval specification factor at metamorphosis results in the Pdm+ neurons' withdrawal from the MB and their redirection to non-MB targets (Truman, 2023).
Another way of recruiting neurons to the larval MB could involve modification of Notch signaling during a GMC division. Data on the DL1 lineage show six examples of GMCs whose division results in one daughter being an MBON and the other being a central complex cell. The actual relationship of MBON-g/LAL.s-NO2i.b to MBON-h/MBON 09 within DAL-v2/3 lineage is not known, but a relationship like that seen in the DL1 lineage is proposed, with one daughter becoming an MBON (08 or 09) and the other a central complex neuron (LAL.s-NO2i.b). In this scenario, the four neurons come from two successive, embryonic-born GMCs. In the larva, however, both siblings become MBONs. This larval similarity could arise from altering embryonic Notch signaling. Typically, the 'Notch-ON' phenotype of the A sibling is established through the Notch target Hey (Hairy/enhancer-of-split like with a Y), a bHLH-O transcription factor Interestingly, in the Kenyon cell lineages, Hey expression is independent of Notch making both siblings 'Notch-ON' and identical. A similar change in Notch state may have occurred for relevant GMCs of the DAL-v2/3 lineage, thereby allowing both siblings to express the MBON fate in the larva. The reestablishment of the normal Notch relationship at metamorphosis might then cause the MBON-gs to lose their MBON characteristic and to acquire their appropriate phenotypes as central complex neurons (Truman, 2023).
The neuronal identity in the insect CNS is generated according to a spatial and temporal pattern that is highly conserved through evolution. The earliest born neurons are often diverse sets of projection neurons that are the basis for the stereotyped tracts and commissures that characterize insect neuropils. Local interneurons are born later in lineages, and these become more similar as the lineage progresses. The evolution of the holometabolous larva involved a shortening of embryonic development, producing a simplified larval body form that could successfully compete for ephemeral food sources. This shortening of insect embryogenesis, though, had a profound impact on their neuroblast-based mode of neurogenesis, resulting in a neurogenic arrest before lineages were complete. Hence, not only do larvae hatch with fewer neurons than found in the mature nervous system, but they should have only the types of neurons characteristic of the early portion of each lineage. Such a truncation may produce mismatches between the neuron types that are needed and those that have been made, i.e., required late-born cell types may be missing and the larva may have early-born cell types that it does not need! Analysis of the developmental relationship of the larval to the adult MB provides insight into how these mismatches are resolved (Truman, 2023).
The relationships of Kenyon cell classes to the MB vertical and medial lobes are highly conserved. The γ Kenyon cells are born first, and their axons form a medial lobe. They are followed by branched, αβ-type neurons, whose vertical axon branch forms a vertical (α) lobe and whose medial, β branch joins the γ cell axons in the medial lobe. In direct developing insects like crickets, both γ and αβ-type Kenyon cells arise during embryogenesis to form the MB of the hatchling cricket, but the hatchling Drosophila has only the early-born γ neurons in its MB. In the absence of late-born αβ-type neurons, the γ neurons are modified with a larval-specific vertical axon branch that provides the basis for the larval vertical lobe. The MBINs and MBONs that provide the input-output circuitry for the larval lobes are neurons that either function in the MB circuit in both larva and adult or function as MB neurons only temporarily in the larva. For the latter cells, it is concluded that their larval phenotype is a transient adaptation for the larval stage and their adult phenotype is more like that seen in their direct-developing ancestors. The deviation of a neuron's larval phenotype from its adult phenotype therefore represents an adaptation to accommodate the lack of needed, late-born neurons (Truman, 2023).
The derived phenotypes of the larval MB neurons relate to their mature phenotypes in the adult brain are summarized (see Each KC type transmits information to multiple compartments.). Previous studies on the abdominal nervous system showed that many larval neurons come from a pool of neurons that die during embryogenesis in direct developing insects, but their death is delayed in metamorphic insects to allow these neurons to function in the larval CNS. In the MB circuit, it was found that the only neurons that were recruited from this pool of 'doomed' neurons were the four larval PAM neurons. The death, though, is perplexing because they are a neuron type needed by the adult, as shown by the addition of ~150 postembryonic born PAM neurons to the adult brain. It is thought that the death of the larval PAM neurons may be related to mechanisms that allow a particular neuron class to be expanded within its lineage. Two clusters of dopamine neurons homologous to the Drosophila PAM clusters are already enlarged in locusts, so this expansion likely occurred before the evolution of complete metamorphosis. Possibly, very ancient insects made only a few PAM-like neurons, and these appeared early in their lineage at an appropriate timing for innervating the early-born γ neurons. If different neuron types maintain their relative order of production within this lineage, then a major expansion of an early-born PAM class would greatly delay the production of later neuron types. A modification that resulted in a PAM phenotype reappearing at the end of the lineage, however, would provide a late-born PAM class that could increase in number without interfering with the appearance of the earlier neurons. Developmental complications of integrating two sets of PAM cells (a small early set and an expanded late set) may have favored the late-born neurons, and thereby necessitated programmed cell death to remove the early-born PAM cells. This scenario brings up the intriguing possibility that after being consigned to the graveyard of unneeded embryonic cells for millions of years, the evolution of the larval stage provided a reason for these neurons to be 'resurrected' as temporary early-born PAM cells for use in the larval CNS (Truman, 2023).
Except for the PAM neurons, embryonic-born neurons that function in the larval MB also function in the adult, either in its MB or in non-MB circuits. The MBINs and most of the MBONs whose terminal role continues in the MB have similar positions in both the larval and the adult MB. For a few MBONs, though, their larval MB function differs markedly from their adult MB function. The embryonic born, glutaminergic MBONs provide informative examples. The increased number of medial lobe compartments in the adult are innervated by more glutaminergic MBONs than are the smaller number of larval medial lobe compartments. The full set of adult glutaminergic MBONs are made during embryogenesis, however. Most take up similar positions in the larval medial lobe while the 'extra' cells are modified to function in the larval vertical lobe. In this way, they temporarily substitute for the late-born cells that normally supply vertical lobe compartments. Many of the neurons that supply the larval-specific compartments of the intermediate peduncle and vertical lobes are neurons fated for non-MB circuits, and modified in the larval for only temporary use in the MB. Many are fated for adult-specific neuropils, such as the central complex or the optic lobes. These temporary, larval MB neurons appear most frequently in lineages that also make permanent MBONs or MBINs. These cells already have the appropriate lineage information to produce the desired phenotypes; they only need to alter the phenotypic read-out of their temporal information and/or Notch state (Truman, 2023).
It is suggested that a hypothetical 'larval specifying factor(s)' is involved in altering the interpretation of the temporal and spatial factors that establish neuronal phenotypes. The expression of such a factor would maintain the larval state but its disappearance at metamorphosis would mean that the larval phenotype could no longer be maintained, and the neurons could change to a mature phenotype appropriate to their temporal and spatial instructions. Indeed, recent studies show that the BTB/POZ transcription factor, Mamo, appears at metamorphosis and is need for the partially dedifferentiated larval neurons to acquire their adult state . While the existence of a larval specification factor is hypothetical in this context, three transcription factors, chinmo, broad, and E93, act as master genes to specify the different life stages of Drosophila . Also, in some insects, the larval stage is actively maintained by the sesquiterpene hormone, juvenile hormone, acting through its major target, the Krüppel-homolog 1 transcription factor. These genes may provide an entry into discovering how the development of terminal fates can be temporarily diverted to produce a novel, larval identity (Truman, 2023).
Animal behavior is often organized into stereotyped sequences that promote the goals of reproduction, development, and survival. However, for most behaviors, the neural mechanisms that govern the order of execution of the motor programs within a sequence are poorly understood. An important model in understanding the hormonal determinants of behavioral sequencing is the ecdysis sequence, which is performed by insects at each developmental transition, or molt. The adult ecdysis sequence in Drosophila includes the emergence of the insect from the pupal case followed by expansion and hardening of the wings. Wing expansion is governed by the hormone bursicon, and stimulation of the bursicon-expressing neurons in newly eclosed flies induces rapid wing expansion. This study shows that that such stimulation delivered prior to eclosion has no immediate effect, but does cause rapid wing expansion after eclosion if the stimulus is delivered within 40 min of that event. A similar delayed effect was observed upon stimulation of a single pair of bursicon-expressing neurons previously identified as command neurons for wing expansion. It is concludes that command neuron stimulation enables the motor output pathway for wing expansion, but that this pathway is blocked prior to eclosion. By manipulating the time of eclosion, this study demonstrates that some physiological process tightly coupled to adult ecdysis releases the block on wing expansion. Eclosion thus serves as a behavioral checkpoint and complements hormonal mechanisms to ensure that wing expansion strictly follows eclosion in the ecdysis sequence (Peabody 2013).
The peptidergic Pigment-dispersing factor (PDF)-Tri neurons are a group of non-clock neurons that appear transiently around the time of adult ecdysis (=eclosion) in the fruit fly Drosophila melanogaster. This specific developmental pattern points to a function of these neurons in eclosion or other processes that are active around pupal-adult transition. As a first step to understand the role of these neurons, the anatomy of the PDF-Tri neurons were characterized. In addition, a further set of peptidergic neurons is described that have been associated with eclosion behavior, eclosion hormone (EH), and crustacean cardioactive peptide (CCAP) neurons, to single cell level in the pharate adult brain. PDF-Tri neurons as well as CCAP neurons co-express a classical transmitter indicated by the occurrence of small clear vesicles in addition to dense-core vesicles containing the peptides. In the tritocerebrum, gnathal ganglion and the superior protocerebrum PDF-Tri neurites contain peptidergic varicosities and both pre- and postsynaptic sites, suggesting that the PDF-Tri neurons represent modulatory rather than pure interneurons that connect the subesophageal zone with the superior protocerebrum. The extensive overlap of PDF-Tri arborizations with neurites of CCAP- and EH-expressing neurons in distinct brain regions provides anatomical evidence for a possible function of the PDF-Tri neurons in eclosion behavior (Selcho, 2018).
Insect mushroom bodies (MB) have an ensemble of synaptic connections well-studied for their role in experience-dependent learning and several higher cognitive functions. MB requires neurotransmission for an efficient flow of information across synapses with different flexibility to meet the demand of the dynamically changing environment of an insect. Neurotransmitter transporters coordinate appropriate changes for an efficient neurotransmission at the synapse. To date, there is no transporter reported for any of the previously known neurotransmitters in the intrinsic neurons of MB. This study reports a highly enriched expression of Choline Transporter (ChT) in Drosophila MB. Knockdown of ChT in a sub-type of MB neurons called alpha/beta core (alpha/betac) and Upsilon neurons leads to eclosion failure, peristaltic defect in larvae, and altered NMJ phenotype. These defects were neither observed on knockdown of proteins of the cholinergic locus in alpha/betac and Upsilon neurons nor by knockdown of ChT in cholinergic neurons. Thus, this study provides insights into non-canonical roles of ChT in MB (Hamid, 2018).
The contribution of circadian clocks in determining time taken for pre-adult development has remained unclear. This study presents results of studies aimed to understand this influence by examining populations of fruit flies carrying three different alleles of the period gene and hence having different free-running periods. Attempts were made to achieve similarity of genetic background among the three strains while also ensuring that they harbored sufficient variation on loci other than period gene. Under constant conditions, flies with long period were found to have slower development whereas in presence of light-dark cycles (LD) of various lengths, the speed of development for each genotype is influenced by whether their eclosion rhythms can entrain to them. Under LD 12:12 (T24), where all three strains entrain, they do not show any difference in time taken for emergence, whereas under LD 10:10 (T20) where long period flies do not entrain and LD 14:14 (T28) where short period flies do not entrain, they have slower and faster pre-adult development, respectively, compared to the controls. A prior stage in development, namely pupation, is not rhythmic, though time taken for pupation is determined by both the environmental cycle and period allele (Srivastava, 2018).
Metamorphosis is an intricate developmental process in which large-scale remodelling of mRNA and microRNA (miRNA) profiles leads to orchestrated tissue remodelling and organogenesis. Whether, which, and how, ribonucleases (RNases) are involved in the RNA profile remodelling during metamorphosis remain unknown. Human Regnase-1 (also known as MCPIP1 and Zc3h12a) RNase remodels RNA profile by cleaving specific RNAs and is a crucial modulator of immune-inflammatory and cellular defence. This study characterized Drosophila CG10889, which was named Drosophila Regnase-1, an ortholog of human Regnase-1. The larva-to-adult metamorphosis in Drosophila includes two major transitions, larva-to-pupa and pupa-to-adult. regnase-1 knockout flies developed until the pupa stage but could not complete pupa-to-adult transition, dying in puparium case. Regnase-1 RNase activity is required for completion of pupa-to-adult transition as transgenic expression of wild-type Drosophila Regnase-1, but not the RNase catalytic-dead mutants, rescued the pupa-to-adult transition in regnase-1 knockout. High-throughput RNA sequencing revealed that regnase-1 knockout flies fail to remodel mRNA and miRNA profiles during the larva-to-pupa transition. Thus, this study has uncovered the roles of Drosophila Regnase-1 in the larva-to-adult metamorphosis and large-scale remodelling of mRNA and miRNA profiles during this metamorphosis process (Zhu, 2019).
Fruit flies (Drosophila melanogaster) eclose from their pupae mainly around dawn. The timing of eclosion is thought to confer adaptive benefits to the organisms and thus shows remarkable accuracy. This study examined the relative contributions of gating of eclosion by the circadian clock versus clock-independent developmental rates and light-induced responses in the eclosion phenotype exhibited by fly populations that have evolved greater accuracy in eclosion rhythms compared to controls. The results showed that stocks that have evolved greater accuracy of eclosion rhythms due to artificial selection do not show reduced individual variation in pupariation and pigmentation time compared to controls, though they do exhibit lower variation in eclosion time. Selected stocks also did not show lower variation in eclosion time under constant light conditions in contrast to the greater accuracy seen under light-dark cycles. Moreover, manipulations of developmental rate by varying larval density and inducing eclosion by changing onset of light phase did not alter the eclosion profile of selected stocks as much as it did controls, since selected stocks largely restricted eclosion to the daytime. These results suggest that fly populations selected for greater accuracy have evolved accurate eclosion rhythms primarily by strengthening circadian gating of eclosion rather than due to fine-tuning of clock-independent developmental processes (Varma, 2019).
Peptidergic neurons are a group of neuronal cells that synthesize and secrete peptides to regulate a variety of biological processes. To identify genes controlling the development and function of peptidergic neurons, a screen was conducted of 545 splice-trap lines and 28 loci were identified that drove expression in peptidergic neurons when crossed to a GFP reporter transgene. Among these lines, an insertion in the alan shepard (shep) gene drove expression specifically in most peptidergic neurons. shep transcripts and SHEP proteins were detected primarily and broadly in the central nervous system (CNS) in embryos, and this expression continued into the adult stage. Loss of shep resulted in late pupal lethality, reduced adult life span, wing expansion defects, uncoordinated adult locomotor activities, rejection of males by virgin females, and reduced neuropil area and reduced levels of multiple pre-synaptic markers throughout the adult CNS. Examination of the bursicon neurons in shep mutant pharate adults revealed smaller somata and fewer axonal branches and boutons, and all of these cellular phenotypes were fully rescued by expression of the most abundant wild-type shep isoform. In contrast to shep mutant animals at the pharate adult stage, shep mutant larvae displayed normal bursicon neuron morphologies. Similarly, shep mutant adults were uncoordinated and weak, while shep mutant larvae displayed largely, though not entirely, normal locomotor behavior. Thus, shep plays an important role in the metamorphic development of many neurons (Chen, 2014).
Peptidergic neurons produce small peptides, called neuropeptides, which are secreted
within the nervous system to influence the activity of other neurons or into the blood to
act on other tissues. Through these targets, neuropeptides regulate a wide range of
processes, which include development, feeding, growth, aggression, reproduction and
learning and memory. One of the first genes identified to play a specific role in the development of peptidergic neurons was dimmed (dimm), which encodes a basic helix-loop-helix transcription factor that is required for the differentiation of diverse peptidergic neurons. Dimm is a key regulator of expression of
the neuropeptide biosynthetic enzyme, peptidylglycine-alphahydroxylating
monooxygenase (Phantom or Phm), and it promotes the differentiation of
neurosecretory properties in many neurons. Both Dimm and
Phm are expressed widely and specifically in peptidergic neurons. In fact, Dimm was
first identified by virtue of its pattern of peptidergic neuron expression through an
enhancer-trap screen. Similar expression pattern-based strategies
may be useful for identification of other factors critical for peptidergic neuron
development (Chen, 2014).
This study sought to identify similar factors through a splice-trap screen for
genes with peptidergic cell-specific expression patterns. 28 insertions were identified with different patterns of peptidergic cell reporter gene expression, driven by P element splicetrap
insertions in specific loci. These insertions drove reporter expression in insulin-like
peptide 2 (ILP2), crustacean cardioactive peptide (CCAP)/bursicon, -RFamide, Furin 1,
and leucokinin (LK) cells and often caused defects typical of disrupted neuropeptide
signaling. Thus, all 28 of these genes are strong candidate regulators of peptidergic cell
development or function (Chen, 2014).
One of the splice-trap insertions was mapped to an exon of the alan shepard (shep) gene, and this insertion was chosen for further analysis because it displayed an expression pattern that was highly similar to Phm and Dimm. shep in situ hybridization and anti-Shep immunostaining later revealed that both the shep mRNA and Shep protein expression is enriched in most neurons, yet shep mutants displayed defects in adult eclosion and wing expansion that suggested specific disruptions in signaling by bursicon and other neuropeptides. Consistent with these behavioral phenotypes, the shep mutant bursicon
neurons had smaller somata, fewer axon branches, and smaller and fewer neuroendocrine
boutons, and all of these phenotypes were rescued by expression of a wild-type shep
cDNA. Interestingly, pan-neuronal RNA interference to shep produced smaller CNS
neuropils and defects in general locomotor behaviors, such as flipping and climbing.
Most of the locomotor phenotypes were restricted to the adult stage, and the effects of
shep mutations on neuronal growth were restricted to pupal development. Thus, shep regulates metamorphic growth of the bursicon neurons, and it may also serve as a general regulator of neuronal growth during metamorphic remodeling (Chen, 2014).
Anti-Shep immunostaining and additional shep reporter genes confirmed
expression in peptidergic neurons, but these markers and shep in situ hybridization also
revealed widespread expression in the CNS, with much lower expression in other tissues.
Shep is orthologous to the c-myc single-strand binding protein, MSSP-2: Previous
studies have described shep as homologous to the vertebrate genes, Rbms2/Scr3
(Armstrong, 2006) or Rbms1/Scr2/MSSP-2. Phylogenetic analysis supports the placement of Shep in the MSSP family, with the
ELAV family of RNA-binding proteins being the next most closely related.
In general, MSSP proteins contain RNA recognition motifs and have been found in
vertebrates to bind DNA, RNA, or proteins to regulate a variety of biological processes,
including DNA polymerization, gene expression, cell transformation, and apoptosis. In Drosophila, Shep interacts with the insulator proteins
Mod(mdg4)2.2 and Su(Hw) to negatively regulate chromosomal insulator activities,
specifically in the CNS (Matzat, 2012). These molecular insights suggest a gene
regulatory mechanism by which Shep may control aspects of the metamorphic
development of the bursicon neurons, as well as other neurons that contribute to the
overall structure of adult brain neuropils (Chen, 2014).
The shep mutant defects in wing expansion presented an opportunity to define
cellular functions of Shep in an experimentally accessible cell type, the bursicon
neurons. In shep mutants, a reduction was observed in the post-pruning growth of the bursicon neurons during metamorphosis, resulting in smaller somata and less branching
in the peripheral axon arbor in pharate adult animals.
Interestingly, the regulation of bursicon neuron growth by shep was stage-dependent. Defects were observed in bursicon neuron soma growth and axon branching during
metamorphosis in hypomorphic shep mutant animals of multiple genotypes, including
shepBG00836/shepBG00836, shepExel6103/shepExel6104 and shepBG00836/shepED210. However, in each of these genotypes, the larval cellular morphologies were
normal.
Other behavioral defects were observed that suggested that the metamorphosis-specific
actions of Shep were not limited to the bursicon neurons. For example, the most severe
shep loss-of-function genotype tested was elav>shep-RNAi, Dicer-2, but
elav>shep-RNAi, Dicer-2 larvae displayed normal crawling distances and self-righting
behaviors, while this genotype showed lethality in the late pupal stages and severe
locomotor defects in adult animals. Associated with this increase during metamorphosis
in the dependence of the nervous system on shep activity, there is also a marked increase
in the levels of shep expression at the onset of metamorphosis (Chen, 2014).
These results provide indirect evidence to suggest that an increase in shep expression during the pupal stage may support neuronal remodeling or other aspects of neuronal
function and development in diverse neurons during metamorphosis.
Although most of the larval behaviors assayed were unaffected in shep mutant animals, one behavioral phenotype in was observed in elav>shep-RNAi, Dicer-2 larvae, namely a
tendency to remain in the center of the apple juice-agarose plate while making many
sharp turns along the path of locomotion. Based on anti-Shep immunostaining, UAS-shep-RNAi, Dicer-2 provided a more complete knockdown of anti-Shep
immunostaining in the CNS than shep RNAi without UAS-Dicer-2 or in shepBG00836
homozygotes or shepBG00836/shepED210 mutant larvae. Moreover, shep RNAi without
UAS-Dicer-2 led to a greater knock-down of Shep in Western blots than
shepExel6103/shepExel6104 (Matzat, 2012). Taken together with the above observation
that many of the weaker shep loss-of-function genotypes had defects that were only
manifest in adults, these findings suggest that shep plays a stage-dependent (largely
metamorphosis-specific) role in the maintenance, function, or development of the
nervous system (Chen, 2014).
The Shep expression pattern and shep
mutant phenotypes reported in this study are consistent with broad actions of this protein in
neuronal development and functions throughout the nervous system. Pan-neuronal loss
of shep resulted in late-pupal lethality and reduced adult life span under both fed and
starved conditions, as well as diverse developmental and behavioral defects, including
failure to complete wing expansion, uncoordinated and weakened adult locomotion,
reduced neuropil areas, and altered mating behaviors. Other groups have also shown
defects in gravitaxis and reduced starvation resistance in shep mutants (Armstrong, 2006; Harbison, 2004; Chen, 2014 and references therein).
Such widespread actions may also explain the partial rescue of the mating defects by
UAS-shep expression in shepBG00836/shepED210 females. Although the possibility cannot be excluded that other Shep isoforms in addition to Shep-E/G (used to create UAS-shep)
str necessary to support the normal function of the post-copulatory grooming circuits, it
is also possible that neurons required for female receptivity to the male may have been
included in the shepBG00836 expression pattern used to drive shep rescue, whereas the
neurons involved in normal post-copulatory grooming behaviors are not (Chen, 2014).
The observation of several seemingly independent behavioral defects (e.g., gravitaxis and
female receptivity to mating) and reduced neuropil areas, taken together with the cellular
defects described in shep-mutant bursicon neurons, suggests that Shep may have
pleiotropic effects on neurite development or other processes throughout the CNS. Such
pleiotropic effects of shep mutations in the CNS may be due to the loss of Shep
suppression of widely distributed chromatin insulator complexes (Matzat, 2012),
so as to establish altered chromatin states and gene expression, potentially in multiple
signaling pathways controlling a range of developmental and physiological events. In
addition, some of the adult shep loss-of-function phenotypes, such as reduced lifespan
and altered mating behaviors, may reflect adult-specific (acute) effects of Shep on
neuronal activity. Alternatively, the metamorphosis-specific regulation of neurite
branching and cell growth in the bursicon neurons may be representative of the actions of
Shep in many neuronal cell types. It will be important in future studies to distinguish
among these models, as the results demonstrate that Shep is a general regulator of the
postembryonic development of mature neurons (Chen, 2014).
In contrast with Drosophila melanogaster, practically nothing is known about the involvement of the TGF-β signaling pathway in the metamorphosis of hemimetabolan insects. To partially fill this gap, the role of Smad factors in the metamorphosis of the German cockroach, Blattella germanica, was studied. In Drosophila, Mad is the canonical R-Smad of the BMP branch of the TGF-β signaling pathway, Smox is the canonical R-Smad of the TGF-β/Activin branch and Medea participates in both branches. In insects, metamorphosis is regulated by the MEKRE93 pathway, which starts with juvenile hormone (JH), whose signal is transduced by Methoprene-tolerant (Met), which stimulates the expression of Kruppel homolog 1 (Kr-h1) that acts to repress E93, the metamorphosis trigger. In B. germanica, metamorphosis is determined at the beginning of the sixth (final) nymphal instar (N6), when JH production ceases, the expression of Kr-h1 declines, and the transcription of E93 begins to increase. The RNAi of Mad, Smox and Medea in N6 of B. germanica reveals that the BMP branch of the TGF-beta signaling pathway regulates adult ecdysis and wing extension, mainly through regulating the expression of Bursicon, whereas the TGF-beta/Activin branch contributes to increasing E93 and decreasing Kr-h1 at the beginning of N6, crucial for triggering adult morphogenesis, as well as to regulating the imaginal molt timing (Santos, 2016).
Animals must adjust their metabolism as they progress through development in order to meet the needs of each stage in the life cycle. This study shows that the dHNF4 nuclear receptor acts at the onset of Drosophila adulthood to direct an essential switch in lipid metabolism. Lipid stores are consumed shortly after metamorphosis but contribute little to energy metabolism. Rather, dHNF4 directs their conversion to very long chain fatty acids and hydrocarbons, which waterproof the animal to preserve fluid homeostasis. Similarly, HNF4alpha is required in mouse hepatocytes for the expression of fatty acid elongases that contribute to a waterproof epidermis, suggesting that this pathway is conserved through evolution. This developmental switch in Drosophila lipid metabolism promotes lifespan and desiccation resistance in adults and suppresses hallmarks of diabetes, including elevated glucose levels and intolerance to dietary sugars. These studies establish dHNF4 as a regulator of the adult metabolic state (Storelli, 2018).
Drosophila breeds and feeds on rotting fruits, which are ephemeral ecosystems. Thus, while the Drosophila larva lives inside a semi-liquid nutritive substrate and feeds extensively to support its rapid increase in mass, the mature adult fly emerges into an unpredictable environment where it uses flight for dispersal and the colonization of more favorable niches. Similar to all insects, the large surface-to-volume ratio of Drosophila adults makes them vulnerable to dehydration. In addition, the metabolic demands of flight require a high rate of gas exchange via the respiratory system, increasing water loss. This study shows that the successful transition from a stationary and protected pupa to a motile adult that can survive in a dry environment is dependent on the dHNF4 nuclear receptor, which acts in the oenocytes to direct a transcriptional program that supports the rapid production of very long-chain fatty acids (VLCFAs) and cuticular hydrocarbons. This activity is consistent with earlier studies, which have demonstrated an important role for oenocytes in VLCFA/hydrocarbon synthesis and desiccation resistance, and places dHNF4 as a central transcriptional regulator for oenocyte function. This study shows, however, that whole-body and hemolymph TAG levels drop to low levels between emergence and day 3 of adulthood, on either a sugar-free diet or a rich 15% sugar diet. The lipids contained in larval adipose cells thus appear to be consumed independently of nutritional input, suggesting that they contribute to non-energetic functions. Moreover, control adults can survive for weeks in the absence of stored lipids, further indicating that they provide a relatively minor source of energy at this stage in development (Storelli, 2018).
It is proposed that lipids in the newly emerged adult are shunted toward a purpose that is more profound than energy production at this stage in development -- the formation of a waterproof cuticle -- and that this occurs in a dHNF4-dependent manner. Stored lipid levels are unaffected in newly emerged dHNF4 mutant adults relative to controls. Lipids, however, accumulate rapidly after this stage in oenocytes, in the presence or absence of nutrients, representing a developmental role for dHNF4 in maintaining lipid homeostasis in this cell type. The timing of lipid accumulation in the oenocytes of starved and fed dHNF4 mutants coincides with the timing of lipid loss and larval adipose cell death. Genetic studies demonstrate that dHNF4 is specifically required in oenocytes to suppress developmental steatosis. This appears to be due to its role in inducing the expression of genes involved in VLCFA elongation. Oenocyte-specific silencing of KAR or ACC recapitulates the developmental steatotic phenotype, while silencing genes that encode downstream or upstream steps in the hydrocarbon biosynthetic pathway has no effect on the lipid levels in oenocytes. Further studies are needed to determine the molecular mechanisms that link VLCFA elongation with oenocyte lipid homeostasis. Finally, adults normally emerge with low hydrocarbon levels that increase dramatically over 3 days, even in the absence of food, in agreement with the hypothesis that the larval adipose cells provide the primary source of precursors for hydrocarbon production. These results are consistent with previous reports, which have shown that dietary manipulation during larval life affects the blend of hydrocarbons in young adult. Consistent with this, oenocyte-specific dHNF4 RNAi in mature adults was shown to lead to a similar degree of lipid accumulation as that seen upon inducing RNAi at emergence. Further studies are required to address this continuing role for dHNF4 in mature adults. In addition, the mechanisms that partition stored lipids toward VLCFA/hydrocarbon production or energy production remain to be determined. Finally, the key functions described for dHNF4 in oenocytes are shared between males and females, indicating that they are not sex specific (Storelli, 2018).
Studies of starvation resistance indicate that newly emerged Drosophila adults can tolerate the absence of nutrients for at least a week. During the non-feeding period of metamorphosis, pupae consume about half of their glycogen and TAG stores and most of the trehalose gathered during the larval stages. In spite of this depletion of energy reserves, sufficient nutrients persist to allow the newly emerged adult to survive and disperse in their new environment. Under hydrated conditions, control lines emerge with the ability to survive from 1 to 3 weeks in the absence of nutrients. It is suggested that these conditions provide insufficient humidity to accurately distinguish the effects of dehydration from the effects of nutrient depletion. Without moisture or food, Drosophila die in less than a day, with a median lifespan of 13 hr. Thus, as is well known in mammals, sufficient hydration is critical for assessing the ability of animals to properly mobilize stored nutrients for survival during starvation (Storelli, 2018).
Changes in metabolism must accompany each stage in development in order to allow normal progression through the life cycle. Although little is known about how development and metabolism are coupled, nuclear receptors appear to play a central role in this process. Consistent with this, a previous study of dHNF4 showed that it is highly up-regulated at the end of metamorphosis as the fly begins its adult life, coordinately inducing genes involved in glucose homeostasis and oxidative phosphorylation (Barry, 2016). Functional studies showed that dHNF4 acts at this stage in the insulin-producing cells to maintain glucose-stimulated insulin secretion and acts in the fat body to promote glucose clearance. This study expands upon these activities for dHNF4 at the onset of adulthood to include an essential role in supporting VLCFA and hydrocarbon production in the oenocytes to allow adult survival and dispersion (Storelli, 2018).
Key genes involved in VLCFA/hydrocarbon production are induced at the onset of adulthood in control adults and reduced in expression in dHNF4 mutants, including KAR, Cpr, Cyp4g1, and several genes encoding predicted elongases. In addition, dHNF4 transcriptional activity can be activated by LCFAs and VLCFAs, which appear to act as ligands for this nuclear receptor. This is consistent with studies of mammalian HNF4α, which have identified LCFAs as ligands that can trigger the conformational changes required for coactivator recruitment. Thus, the free fatty acids generated by lipolysis from the larval adipose cells could act as ligands for dHNF4 in oenocytes, driving the transcription of genes in the VLCFA/hydrocarbon pathway. The resulting VLCFAs might further activate dHNF4, providing a feed-forward loop to enhance VLCFA/hydrocarbon production. These levels of regulation could ensure that free fatty acids are rapidly and efficiently converted into VLCFAs and hydrocarbons to provide an effective waterproof barrier for the young adult (Storelli, 2018).
Simple carbohydrates are required for optimal Drosophila longevity, yet dHNF4 and VLCFA/hydrocarbon biosynthesis are required in oenocytes to provide tolerance to these nutrients. In addition, increased humidity is sufficient to rescue sugar toxicity and hyperglycemia in dHNF4 mutants, suggesting that defects in fluid homeostasis are contributing to these phenotypes. Defects in fluid homeostasis could occur in dHNF4 mutants because of impaired waterproofing of the cuticle or the trachea. Parvy and colleagues demonstrated that, besides providing precursors for hydrocarbon synthesis, VLCFA metabolism in oenocytes is required to remotely waterproof the respiratory system in larvae. Notably, disrupting VLCFA production in oenocytes via ACC, KAR, or FASNCG17374 RNAi in this cell type results in defects in tracheal waterproofing and larval lethality. Lethality between larval and pupal stages was observed when these RNAi constructs were driven with a constitutive oenocyte-specific GAL4 driver. However, defects in tracheal air filling or lethality are not seen in dHNF4 mutant larvae or in animals with constitutive dHNF4 RNAi expression in oenocytes, suggesting that there are no significant effects on tracheal waterproofing in these animals. In addition, altering cuticular hydrocarbon production while leaving VLCFA synthesis intact (by silencing Cyp4g1 in oenocytes) induces sugar toxicity and defects in glucose homeostasis. Thus it is postulated that reduced hydrocarbon production by oenocytes and altered cuticular waterproofing play central roles in the physiological defects observed in dHNF4 mutants (Storelli, 2018).
Understanding of diabetes in humans provides possible models to explain the physiological mechanisms that link dHNF4 activity in oenocytes to fluid and carbohydrate homeostasis. In mammals, excess blood glucose cannot be effectively reabsorbed by the kidney and is excreted with urine. One possibility is that dHNF4 mutants provide a sensitized genetic context for the development of a hyperosmolar hyperglycemic state in Drosophila. Increased transepidermal water loss could reduce hemolymph volume, contributing to the development of hyperglycemia on a sugar-containing diet. This hypothesis could explain the elevated glucose levels observed in dHNF4 mutants and the normalization of glucose levels in high humidity. Further studies are required to test this possibility and dissect the mechanisms that couple dHNF4 function in oenocytes to systemic carbohydrate homeostasis (Storelli, 2018).
A number of studies have focused on characterizing the signaling pathways that govern glucose sensing and metabolism in Drosophila. These studies have provided evidence for glucose-induced cellular damage via oxidative stress or the formation of advanced glycation end-products. In spite of these efforts, however, more remains to be learned about the mechanisms underlying dietary sugar toxicity. This work suggests unexpected roles for oenocyte lipid metabolism and hydration status in regulating glucose homeostasis and suppressing sugar toxicity in Drosophila (Storelli, 2018).
Interestingly, the role of VLCFAs in reducing trans-epidermal water loss is conserved through evolution. Similarly, mouse mutants for Elovl1 and Elovl4 are born normally but die shortly thereafter from acute dehydration, similar to dHNF4 mutant adults. This lethality is accompanied by reduced epithelial barrier function, defects in the lamellar structure of newborn skin, reduced levels of VLCFAs, and the absence of key acylceramides that contribute to skin hydrophobicity. Genetic studies of the gene encoding ELOVL3, which elongates C20 fatty acids to C22 and C24, revealed similar functions. Elovl3 mutant mice have disrupted skin and hair morphology resulting in increased trans-epidermal water loss. Taken together, these results indicate that VLCFA production is critical for proper lipid metabolism and survival after birth in mouse, revealing a perinatal transition that parallels the role for VLCFAs and hydrocarbons in newly emerged Drosophila adults. Interestingly, these studies also link Elovl gene expression to HNF4α function in mice. Elovl3, Elovl5, and KAR are all expressed at reduced levels in HNF4α mutant hepatocytes, indicating that this regulatory link is conserved through evolution. Further studies are required to determine if HNF4α exerts a similar role in mammalian skin and if its activity is required for epithelial barrier function and desiccation resistance (Storelli, 2018).
Tissue remodeling is a crucial process in animal development and disease progression. Coordinately controlled by the two main insect hormones, juvenile hormone (JH) and 20-hydroxyecdysone (20E), tissues are remodeled context-specifically during insect metamorphosis. Previous work has discovered that two matrix metalloproteinases (Mmps) cooperatively induce fat body cell dissociation in Drosophila. However, the molecular events involved in this Mmps-mediated dissociation are unclear. This study reports that JH and 20E coordinately and precisely control the developmental timing of Mmps-induced fat body cell dissociation. During the larval-prepupal transition, the anti-metamorphic factor Kr-h1 was found to transduce JH signaling, which directly inhibited Mmps expression and activated expression of tissue inhibitor of metalloproteinases (timp), and thereby suppressed Mmps-induced fat body cell dissociation. It is also noted that upon a decline in the JH titer, a prepupal peak of 20E suppresses Mmps-induced fat body cell dissociation through the 20E primary-response genes, E75 and Blimp-1, which inhibited expression of the nuclear receptor and competence factor βftz-F1. Moreover, upon a decline in the 20E titer, βftz-F1 expression was induced by the 20E early-late response gene DHR3, and then βftz-F1 directly activated Mmps expression and inhibited timp expression, causing Mmps-induced fat body cell dissociation during 6-12 hrs after puparium formation. In conclusion, coordinated signaling via JH and 20E finely tunes the developmental timing of Mmps-induced fat body cell dissociation. These findings shed critical light on hormonal regulation of insect metamorphosis (Jia, 2017).
MMPs and tissue inhibitor of metalloproteinases (TIMPs) play crucial roles in regulating tissue remodeling in both vertebrates and Drosophila. Previous work has demonstrated the collaborative functions of Mmp1 and Mmp2 in inducing fat body cell dissociation in Drosophila. timp mutant adults show autolyzed tissue in the abdominal cavity and inflated wings, a phenotype consistent with the role of timp in BM integrity and remodeling. The current study clarified the role of timp in inhibiting the enzymatic activity of Mmps and thus, Mmp-induced fat body cell dissociation. In mammals, Mmps activity in vivo is controlled at different levels, including the regulation by gene expression, the zymogens activation, and the inhibition of active enzymes by TIMPs. These studies unify the important inhibitory roles of timp/TIMP in regulating tissue remodeling in both Drosophila and mammals. In addition to regulating Mmp expression, JH and 20E signals differentially regulate timp expression, with the stimulatory role of Kr-h1 and the inhibitory role of βftz-F1. Because timp inhibits the enzymatic activity of Mmps in the Drosophila fat body, it is concluded that JH and 20E coordinately control Mmps activity at both the mRNA and enzymatic levels (Jia, 2017).
Previously work has show the requirement of both JH and its receptors to inhibit fat body cell dissociation in Drosophila. This study demonstrated the ability of Kr-h1 to transduce JH signaling to decrease Mmp expression and to induce timp expression during larval-prepupal transition. Moreover, a Kr-h1-binding sites (KBS) was identified in the Mmp1 promoter, indicating that Kr-h1 directly represses Mmp1 expression. Interestingly, Kr-h1 expression gradually increases from initiation of wandering (IW) to 3 h APF when induced by JH and 20E in an overlapping manner, thus inhibiting the enzymatic activity of Mmps and Mmp-induced fat body cell dissociation during the larval-prepupal transition. Moreover, Kr-h1 acts as an anti-metamorphic factor by inhibiting 20E signaling. It is proposed, in addition to directly affecting the expression of Mmps and timp, that Kr-h1 might also indirectly regulate their expression by inhibiting 20E signaling (Jia, 2017).
Two consecutive 20E pulses control timely metamorphosis in Drosophila. Together with previous findings, the current results show that the conserved 20E transcriptional cascade precisely controls the timing of Mmp-induced fat body cell dissociation. In general, the first 20E signal pulse plays an inhibitory role during the larval-prepupal transition; however, it is a prerequisite for the expression of βftz-F1, which induces the second 20E signal pulse during the prepupal-pupal transition and the expression of Mmps. Because of the requirement for the first 20E signal pulse, blockade of the 20E receptor prevents fat body cell dissociation. When JH titer declines, the prepupal peak of 20E activates expression of two 20E primary-response genes, E75 and Blimp-1, to inhibit fat body cell dissociation: E75 represses DHR3 transactivation of βftz-F1 expression, and Blimp-1 directly represses βftz-F1 expression. During the prepupal-pupal transition, DHR3 directly induces βftz-F1 expression from 6 h APF to 12 APF. Before pupation, βftz-F1 induces Mmp expression and represses timp expression. Moreover, an FBS was identified in the Mmp2 promoter, demonstrating that βftz-F1 directly induces Mmp2 expression. Finally, within 6 h before pupation, Mmp1 and Mmp2 cooperatively induce fat body cell dissociation, with each assuming a distinct role (Jia, 2017).
Insect metamorphosis is coordinately controlled by JH and 20E, whereas the hormonal control of tissue remodeling is strictly context-specific. Different larval tissues and adult organs might have distinct, yet precise, developmental fates and timing. Knowledge regarding this question is poor. Based on previous preliminary information, this study clarified the detailed molecular mechanisms by which JH and 20E precisely control the developmental timing of Mmp-induced fat body cell dissociation at both mRNA and enzymatic levels in Drosophila, and a working model is provided of hormonal control of tissue remodeling in animals (see Model showing developmental timing of Mmp-induced fat body cell dissociation is coordinately and precisely controlled by JH and 20E in Drosophila). In summary, at first, Kr-h1 transduces JH signaling to inhibit Mmp-induced fat body cell dissociation during larval-prepupal transition. Then when JH titer declines, the prepupal peak of 20E suppresses Mmp-induced fat body cell dissociation through E75 and Blimp-1, which inhibit βftz-F1 expression. Finally, until 20E titer declines, DHR3 induces βftz-F1 expression, and βftz-F1 covers the 20E-triggered transcriptional cascade to activate Mmp-induced fat body cell dissociation within 6 h before pupation. This study provides an excellent sample for better understanding the hormonal regulation of insect metamorphosis (Jia, 2017).
Ecdysteroids, typified by 20-hydroxyecdysone (20E), are essential hormones for the development, reproduction and physiology of insects and other arthropods. For over half a century, Drosophila has been used as a model of ecdysteroid biology. Many aspects of the biosynthesis and regulation of ecdysteroids in this species are understood at the molecular level, particularly with respect to their secretion from the prothoracic gland (PG) cells of the ring gland, widely considered the dominant biosynthetic tissue during development. Discrete pulses of 20E orchestrate transitions during the D. melanogaster life cycle, the sources of which are generally well understood, apart from the large 20E pulse at the onset of pharate adult development, which has received little recent attention. As the source of this pharate adult pulse (PAP) is a curious blind spot in Drosophila endocrinology, this study evaluated published biochemical and genetic data as they pertain to three hypotheses for the source of PAP 20E: the PG; an alternative biosynthetic tissue; or the recycling of stored 20E. Based on multiple lines of evidence, it is contended the PAP cannot be derived from biosynthesis, with other data consistent with D. melanogaster able to recycle ecdysteroids before and during metamorphosis. Published data also suggest the PAP is conserved across Diptera, with evidence for pupal-adult ecdysteroid recycling occurring in other cyclorrhaphan flies. Further experimental work is required to test the ecdysteroid recycling hypothesis, which would establish fundamental knowledge of the function, regulation, and evolution of metamorphic hormones in dipterans and other insects (Scanlan, 2023).
Alvarez-Saavedra, E. and Horvitz, H. R. (2010). Many families of C. elegans microRNAs are not essential for development or viability. Curr Biol 20: 367-373. PubMed ID: 20096582
Ashburner, M. (1989). Drosophila: A laboratory handbook. Cold Spring Harbor Laboratory Press, Plainview NY.
Athilingam, T., Parihar, S. S., Bhattacharya, R., Rizvi, M. S., Kumar, A. and Sinha, P. (2022). Proximate larval epidermal cell layer generates forces for Pupal thorax closure in Drosophila. Genetics 221(1). PubMed ID: 35166774
Boulan, L., Martin, D. and Milan, M. (2013). bantam miRNA promotes systemic growth by connecting insulin signaling and ecdysone production. Curr Biol 23: 473-478. PubMed ID: 23477723
Barry, W. E. and Thummel, C. S. (2016). The Drosophila HNF4 nuclear receptor promotes glucose-stimulated insulin secretion and mitochondrial function in adults. Elife 5. PubMed ID: 27185732
Bu, S., Lau, S. S. Y., Yong, W. L., Zhang, H., Thiagarajan, S., Bashirullah, A. and Yu, F. (2023). Polycomb group genes are required for neuronal pruning in Drosophila. BMC Biol 21(1): 33. PubMed ID: 36793038
Companys, P. P., Norris, A. and Bischoff, M. (2020). Coordination of cytoskeletal dynamics and cell behaviour during Drosophila abdominal morphogenesis. J Cell Sci 133(6). PubMed ID: 32229579
Chen, D., Qu, C. and Hewes, R. S. (2014). Neuronal remodeling during metamorphosis is regulated by the alan shepard (shep) gene in Drosophila melanogaster. Genetics [Epub ahead of print]. PubMed ID: 24931409
Colombani, J., et al. (2005). Antagonistic actions of ecdysone and insulins determine final size in Drosophila. Science 310(5748): 667-70. 16179433
Diao, F., Ironfield, H., Luan, H., Diao, F., Shropshire, W. C., Ewer, J., Marr, E., Potter, C. J., Landgraf, M. and White, B. H. (2015). Plug-and-play genetic access to drosophila cell types using exchangeable exon cassettes. Cell Rep 10(8): 1410-1421. PubMed ID: www.ncbi.nlm.nih.gov/pubmed/25732830">25732830 Diao, F., Elliott, A. D., Diao, F., Shah, S. and White, B. H. (2017). Neuromodulatory connectivity defines the structure of a behavioral neural network. Elife 6. PubMed ID: 29165248
Fristrom, D. and Fristrom, J.W. (1993). The metamorphic development of the adult epidermis. In: The Development of Drosophila melanogaster. pp 843-897. Cold Spring Harbor Laboratory Press, Plainview NY. Hamid, R., Hajirnis, N., Kushwaha, S., Saleem, S., Kumar, V. and Mishra, R. K. (2018). Drosophila Choline transporter non-canonically regulates pupal eclosion and NMJ integrity through a neuronal subset of mushroom body. Dev Biol. PubMed ID: 30529058
Hanna, L., Lamouret, T., Pocas, G. M., Mirth, C. K., Moczek, A. P., Nijhout, F. H. and Abouheif, E. (2022). Evaluating old truths: Final adult size in holometabolous insects is set by the end of larval development. J Exp Zool B Mol Dev Evol. PubMed ID: 35676886
Hao, S., Gestrich, J. Y., Zhang, X., Xu, M., Wang, X., Liu, L. and Wei, H. (2021). Neurotransmitters Affect Larval Development by Regulating the Activity of Prothoracicotropic Hormone-Releasing Neurons in Drosophila melanogaster. Front Neurosci 15: 653858. PubMed ID: 34975366
Hartenstein, V. (1993). Atlas of Drosophila development. Cold Spring Harbor Laboratory Press, Plainview, NY.
Hirschhauser, A., Molitor, D., Salinas, G., Grosshans, J., Rust, K., Bogdan, S. (2023). Single-cell transcriptomics identifies new blood cell populations in Drosophila released at the onset of metamorphosis. Development, 150(18) PubMed ID: . PubMed ID: 37681301
Jang, S., Chen, J., Choi, J., Lim, S. Y., Song, H., Choi, H., Kwon, H. W., Choi, M. S. and Kwon, J. Y. (2021). Spatiotemporal organization of enteroendocrine peptide expression in Drosophila. J Neurogenet: 1-12. PubMed ID: 34670462
Jia, Q., Liu, S., Wen, D., Cheng, Y., Bendena, W. G., Wang, J. and Li, S. (2017). Juvenile hormone and 20-hydroxyecdysone coordinately control the developmental timing of matrix metalloproteinase-induced fat body cell dissociation. J Biol Chem 292(52): 21504-21516. PubMed ID: 29118190
Juarez-Carreno, S., Geissmann, F. (2023). The macrophage genetic cassette inr/dtor/pvf2 is a nutritional status checkpoint for developmental timing. Sci Adv, 9(38):eadh0589 PubMed ID: 37729406
Juhász, G., Puskás, L. G., Komonyi, O, Érdi, B., Maróy, P., Neufeld, T. P. and Sass, M. (2007). Gene expression profiling identifies FKBP39 as an inhibitor of autophagy in larval Drosophila fat body. Cell Death Differ. 14: 1181-1190. PubMed Citation: 17363962
Kim, D. H., Han, M. R., Lee, G., Lee, S. S., Kim, Y. J. and Adams, M. E. (2015). Rescheduling behavioral subunits of a fixed action pattern by genetic manipulation of peptidergic signaling. PLoS Genet 11: e1005513. PubMed ID: 26401953
Kim Y-J, Zitnan D, Galizia CG, Cho K-H, Adams ME (2006a). A command chemical triggers an innate behavior by sequential activation of multiple peptidergic ensembles. Current Biology. 16: 1395-1407. PubMed ID: 16860738
Kim Y-J, Zitnan D, Cho K-H, Schooley DA, Mizoguchi A, Adams ME (2006b). Central peptidergic ensembles associated with organization of an innate behavior. Proc Natl Acad Sci USA. 103: 14211-14216. PubMed ID: 16968777
Kozlova, T. and Thummel, C. S. (2002). Spatial patterns of ecdysteroid receptor activation during the onset of Drosophila metamorphosis. Development 129: 1739-1750. 11923209
Lam, G., Nam, H. J., Velentzas, P. D., Baehrecke, E. H. and Thummel, C. S. (2021).
Drosophila E93 promotes adult development and suppresses larval responses to ecdysone during metamorphosis. Dev Biol. PubMed ID: 34648816.
Liu, Y., et al. (2009). Juvenile hormone counteracts the bHLH-PAS transcription factors MET and GCE to prevent caspase-dependent programmed cell death in Drosophila. Development 136(12): 2015-25. PubMed Citation: 19465595
Mark, B., Bustos-Gonzalez, L., Cascallares, G., Conejera, F. and Ewer, J. (2021). The circadian clock gates Drosophila adult emergence by controlling the timecourse of metamorphosis. Proc Natl Acad Sci U S A 118(27). PubMed ID: 34183412
Murakawa, T., Nakamura, T., Kawaguchi, K., Murayama, F., Zhao, N., Stasevich, T. J., Kimura, H. and Fujita, N. (2022). A Drosophila toolkit for HA-tagged proteins unveils a block in autophagy flux in the last instar larval fat body. Development 149(6). PubMed ID: 35319746
Moore, M. C., Taylor, D. T. (2023). Effects of valproate on seizure-like activity in Drosophila melanogaster with a knockdown of Ube3a in different neuronal populations as a model of Angelman Syndrome. Epilepsy & behavior reports, 24:100622 PubMed ID: 37842098
Ninov, N., Chiarelli, D. A. and Martín-Blanco, E. (2007). Extrinsic and intrinsic mechanisms directing epithelial cell sheet replacement during Drosophila metamorphosis. Development 134(2): 367-79. PubMed Citation: 17166923
Nishimura, T. (2020). Feedforward Regulation of Glucose Metabolism by Steroid Hormones Drives a Developmental Transition in Drosophila. Curr Biol. PubMed ID: 32679096
Niwa, R., Niimi, T., Honda, N., Yoshiyama, M., Itoyama, K., Kataoka, H. and Shinoda, T. (2008). Juvenile hormone acid O-methyltransferase in Drosophila melanogaster. Insect Biochem. Mol. Biol. 38: 714-720. PubMed Citation: 18549957
Nunes, C., Koyama, T. and Sucena, E. (2021). Co-option of immune effectors by the hormonal signalling system triggering metamorphosis in Drosophila melanogaster. PLoS Genet 17(11): e1009916. PubMed ID: 34843450
Ohhara, Y., Shimada-Niwa, Y., Niwa, R., Kayashima, Y., Hayashi, Y., Akagi, K., Ueda, H., Yamakawa-Kobayashi, K. and Kobayashi, S. (2015). Autocrine regulation of ecdysone synthesis by β3-octopamine receptor in the prothoracic gland is essential for Drosophila metamorphosis. Proc Natl Acad Sci USA [Epub ahead of print]. PubMed ID: 25605909
Park, Y., Kim, Y. J. and Adams, M. E. (2002). Identification of G protein-coupled receptors for Drosophila PRXamide peptides, CCAP, corazonin, and AKH supports a theory of ligand-receptor coevolution. Proc Natl Acad Sci U S A 99: 11423-11428. PubMed ID: 12177421
Parvy, J. P., Napal, L., Rubin, T., Poidevin, M., Perrin, L., Wicker-Thomas, C. and Montagne, J. (2012). Drosophila melanogaster Acetyl-CoA-carboxylase sustains a fatty acid-dependent remote signal to waterproof the respiratory system. PLoS Genet 8(8): e1002925. PubMed ID: 22956916
Peabody, N. C. and White, B. H. (2013). Eclosion gates progression of the adult ecdysis sequence of Drosophila. J Exp Biol 216(Pt 23):4395-402. PubMed ID: 24031052
Qian, W., Guo, M., Peng, J., Zhao, T., Li, Z., Yang, Y., Li, H., Zhang, X., King-Jones, K. and Cheng, D. (2023). Decapentaplegic retards lipolysis during metamorphosis in Bombyx mori and Drosophila melanogaster. Insect Biochem Mol Biol 155: 103928. PubMed ID: 36870515
Roller L, Zitnanová I, Dai L, Simo L, Park Y, Satake H, et al. Ecdysis triggering hormone signaling in arthropods. Peptides. 31: 429-441. PubMed ID: 19951734
Santos, C. G., Fernandez-Nicolas, A. and Belles, X. (2016). Smads and insect hemimetabolan metamorphosis. Dev Biol [Epub ahead of print]. PubMed ID: 27452629
Sarraf-Zadeh, L., Christen, S., Sauer, U., Cognigni, P., Miguel-Aliaga, I., Stocker, H., Kohler, K. and Hafen, E. (2013). Local requirement of the Drosophila insulin binding protein Imp-L2 in coordinating developmental progression with nutritional conditions. Dev Biol 381: 97-106. PubMed ID: 23773803
Scanlan, J. L., Robin, C. and Mirth, C. K. (2023). Rethinking the ecdysteroid source during Drosophila pupal-adult development. Insect Biochem Mol Biol 152: 103891. PubMed ID: 36481381
Sekyrova, P., Bohmann, D., Jindra, M. and Uhlirova, M. (2010). Interaction between Drosophila bZIP proteins Atf3 and Jun prevents replacement of epithelial cells during metamorphosis. Development 137(1): 141-50. PubMed Citation: 20023169
Selcho, M., Muhlbauer, B., Hensgen, R., Shiga, S., Wegener, C. and Yasuyama, K. (2018). Anatomical characterization of PDF-tri neurons and peptidergic neurons associated with eclosion behavior in Drosophila. J Comp Neurol. PubMed ID: 29427506
Srivastava, M., James, A., Varma, V., Sharma, V. K. and Sheeba, V. (2018). Environmental cycles regulate development time via circadian clock mediated gating of adult emergence. BMC Dev Biol 18(1): 21. PubMed ID: 30577765
Storelli, G., Nam, H. J., Simcox, J., Villanueva, C. J. and Thummel, C. S. (2018). Drosophila HNF4 directs a switch in lipid metabolism that supports the transition to adulthood. Dev Cell 48(2):200-214. PubMed ID: 30554999
Sullivan, A. A. and Thummel, C. S. (2003). Temporal profiles of nuclear receptor gene expression reveal coordinate transcriptional responses during Drosophila development. Mol. Endocrinol. 17(11): 2125-37. 12881508
Tajiri, R., Ogawa, N., Fujiwara, H. and Kojima, T. (2017). Mechanical control of whole body shape by a single cuticular protein Obstructor-E in Drosophila melanogaster. PLoS Genet 13(1): e1006548. PubMed ID: 28076349
Tran, H. T., Cho, E., Jeong, S., Jeong, E. B., Lee, H. S., Jeong, S. Y., Hwang, J. S. and Kim, E. Y. (2018). Makorin 1 regulates developmental timing in Drosophila. Mol Cells. PubMed ID: 30396233
Truman, J. W. and Riddiford, L. M. (2022). Chinmo is the larval member of the molecular trinity that directs Drosophila metamorphosis. Proc Natl Acad Sci U S A 119(15): e2201071119. PubMed ID: 35377802
Truman, J. W., Price, J., Miyares, R. L. and Lee, T. (2023). Metamorphosis of memory circuits in Drosophila reveals a strategy for evolving a larval brain. Elife 12. PubMed ID: 36695420
Vandersmissen, H. P., Hiel, M. B., Loy, T. V., Vleugels, R. and Broeck, J. V. (2014). Silencing D. melanogaster lgr1 impairs transition from larval to pupal stage. Gen Comp Endocrinol [Epub ahead of print]. PubMed ID: 25157788
Varma, V., Krishna, S., Srivastava, M., Sharma, V. K. and Sheeba, V. (2019). Accuracy of fruit-fly eclosion rhythms evolves by strengthening circadian gating rather than developmental fine-tuning. Biol Open 8(8). PubMed ID: 31455663
Yamanaka, N., Marques, G. and O'Connor, M. B. (2015). Vesicle-mediated steroid hormone secretion in Drosophila melanogaster. Cell 163: 907-919. PubMed ID: 26544939
Xie, X. J., et al. (2015). CDK8-Cyclin C mediates nutritional regulation of developmental transitions through the Ecdysone receptor in Drosophila. PLoS Biol 13: e1002207. PubMed ID: 26222308
Zeng, J., Huynh, N., Phelps, B. and King-Jones, K. (2020). Snail synchronizes endocycling in a TOR-dependent manner to coordinate entry and escape from endoreplication pausing during the Drosophila critical weight checkpoint. PLoS Biol 18(2): e3000609. PubMed ID: 32097403
Zhou, X. and Riddiford, L. M. (2002). Broad specifies pupal development and mediates the 'status quo' action of juvenile hormone on the pupal-adult transformation in Drosophila and Manduca. Development 129: 2259-2269. 11959833
Zhu, L., Liao, S. E. and Fukunaga, R. (2019). Drosophila Regnase-1 RNase is required for mRNA and miRNA profile remodelling during larva-to-adult metamorphosis. RNA Biol: 1-15. PubMed ID: 31195914
The Interactive Fly resides on the
Society for Developmental Biology's Web server.
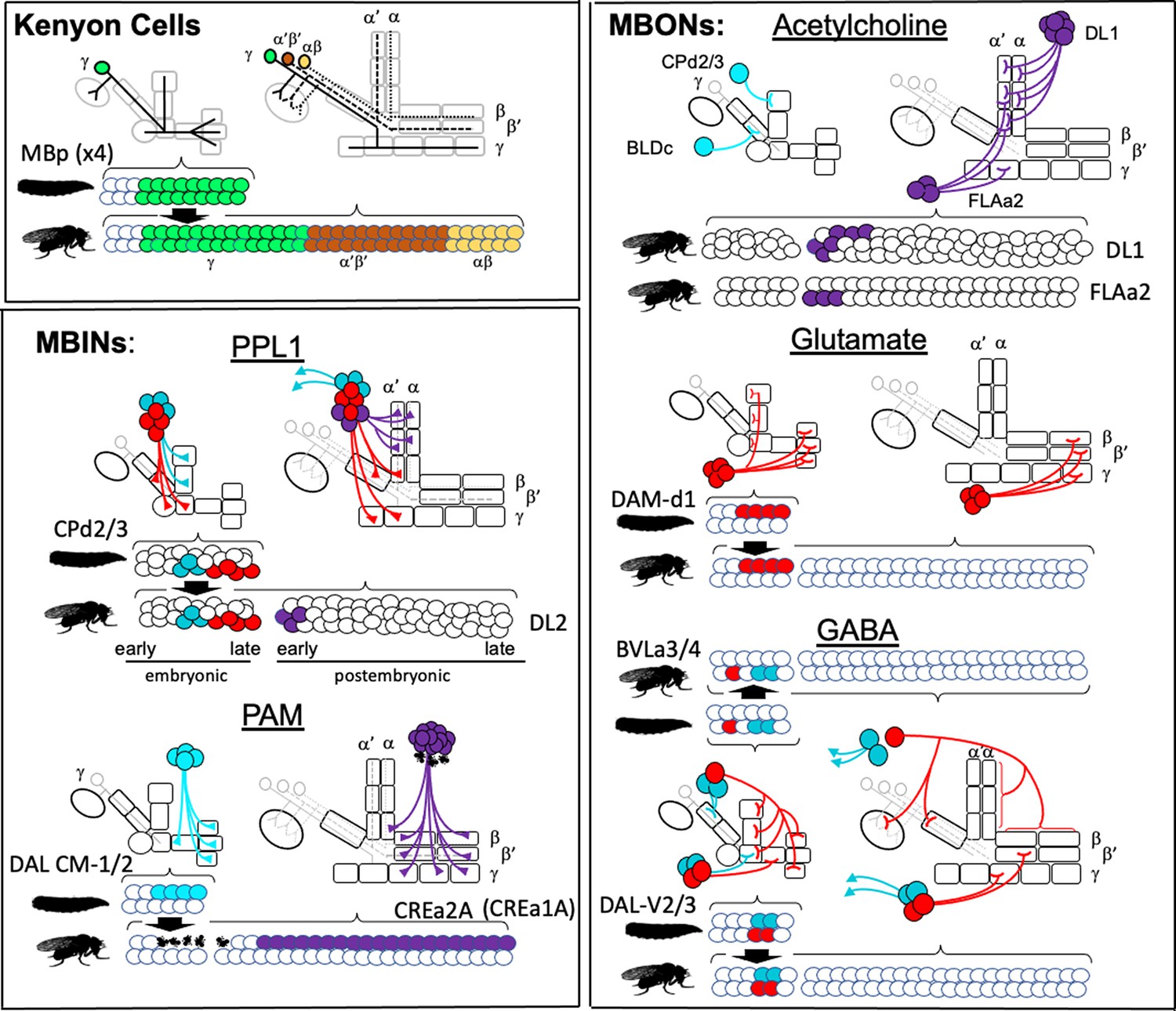{kind=link}